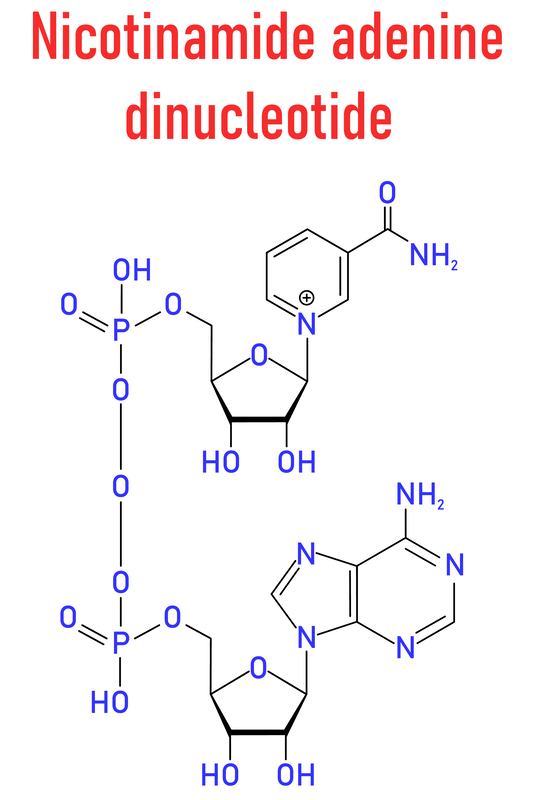
Nicotinamide adenine dinucleotide, or NAD, is a crucial coenzyme found in every cell of our bodies. It plays a fundamental role in various cellular processes, making it a subject of increasing interest in the world of health and wellness. In this blog post, we’ll explore the pros and cons of NAD supplementation, how to read product labels, recommended dosages, best sources, efficacy, potential side effects, and toxicity concerns.
Pros of NAD:
Energy Production: NAD is a key player in the process of converting food into cellular energy through cellular respiration. Its presence can boost your overall energy levels.
Cellular Repair: NAD is essential for DNA repair, promoting healthy aging, and reducing the risk of age-related diseases.
Brain Function: Some studies suggest that NAD may enhance cognitive function and support brain health.
Metabolic Health: NAD is involved in regulating metabolism, which can help with weight management and overall metabolic health.
Anti-Aging: NAD has been linked to longevity and the activation of certain proteins associated with aging.
Cons of NAD:
Limited Absorption: Oral NAD supplements may have limited bioavailability, meaning that not all of the NAD is effectively absorbed by the body.
Expensive: NAD supplements can be costly, especially if you require high doses.
Uncertain Long-Term Effects: While NAD shows promise in various health areas, more research is needed to understand its long-term safety and efficacy fully.
How to Read the Label:
When shopping for NAD supplements, keep the following in mind:
Type: NAD supplements are often labeled as “NAD+” or “Nicotinamide Riboside (NR).” Look for products that specify the type of NAD compound used.
Dosage: Review the recommended dosage on the label, and consider consulting a healthcare professional for personalized guidance.
Dosage:
The appropriate NAD dosage can vary widely depending on individual needs and the specific form of NAD used. Typical daily doses range from 100 mg to 500 mg. However, it’s essential to consult with a healthcare provider to determine the right dosage for your specific health goals.
Best Sources of NAD:
Nicotinamide Riboside (NR): NR is a precursor to NAD and is found in some dietary supplements. It is known for its potential bioavailability and ability to increase NAD levels.
Whole Foods: Foods like meat, fish, dairy, and certain vegetables contain NAD precursors and can support NAD production in the body.
Efficacy:
The efficacy of NAD supplements depends on various factors, including individual health status, age, and the specific form of NAD used. Some studies suggest that NR supplements can effectively raise NAD levels, but more research is needed to establish their long-term benefits fully.
Side Effects:
NAD supplements are generally well-tolerated, with few reported side effects. Some individuals may experience minor gastrointestinal discomfort, headache, or dizziness. If you experience severe or persistent side effects, consult a healthcare provider.
Toxicity:
NAD supplements are not associated with toxicity when taken at recommended doses. However, high doses of NAD may lead to side effects, such as flushing or liver enzyme elevation. It’s crucial to adhere to recommended dosages and seek professional guidance when considering higher doses.
References:
Belenky P, Bogan KL, Brenner C. NAD+ metabolism in health and disease. Trends Biochem Sci. 2007;32(1):12-19.
Trammell SA, Schmidt MS, Weidemann BJ, et al. Nicotinamide riboside is uniquely and orally bioavailable in mice and humans. Nat Commun. 2016;7:12948.
Gomes AP, Price NL, Ling AJ, et al. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013;155(7):1624-1638.
Conclusion:
NAD supplementation holds promise in various health areas, but it’s essential to weigh the pros and cons carefully. While NAD can support cellular energy production, DNA repair, and overall health, its effectiveness may vary among individuals. To make the most of NAD supplementation, consult with a healthcare professional, choose reliable products, and follow recommended dosages. Additionally, stay informed about ongoing research to better understand the long-term benefits and potential risks associated with NAD.
Nicotinamide adenine dinucleotide (NAD) is a coenzyme that plays a pivotal role in cellular energy metabolism, DNA repair, and various other essential cellular processes. Recent research has illuminated the complex relationship between NAD and cancer cells, shedding light on how this molecule can impact cancer development, progression, and treatment. In this blog post, we will explore the fascinating connection between NAD and cancer cells, supported by scientific references.
Understanding NAD:
Before delving into the relationship between NAD and cancer, let’s grasp the basics of NAD:
NAD exists in two forms: NAD+ (oxidized) and NADH (reduced).
NAD is essential for transferring electrons in cellular redox reactions, contributing to energy production.
NAD participates in DNA repair mechanisms and regulates various cellular processes.
NAD and Cancer Cells:
DNA Repair and PARP Inhibition:
NAD’s role in DNA repair is central to its influence on cancer cells. Poly(ADP-ribose) polymerases (PARPs) are enzymes that use NAD+ as a substrate during DNA repair processes. PARP inhibitors have emerged as a promising therapeutic approach in cancer treatment. These inhibitors interfere with DNA repair mechanisms, causing synthetic lethality in cancer cells with impaired DNA repair pathways, such as those with BRCA mutations. PARP inhibitors deplete NAD+ and prevent cancer cells from effectively repairing DNA damage.
References:
Lord, C. J., & Ashworth, A. (2017). PARP inhibitors: Synthetic lethality in the clinic. Science, 355(6330), 1152-1158. doi:10.1126/science.aam7344
NAD and Metabolic Regulation:
Cancer cells often display altered metabolic profiles, and NAD metabolism can play a role in these changes. Research suggests that cancer cells may have increased NAD+ levels, which can promote cell survival and proliferation. Targeting NAD+ metabolism in cancer cells is an active area of investigation. Inhibiting enzymes involved in NAD+ synthesis, such as nicotinamide phosphoribosyltransferase (NAMPT), has shown potential in limiting cancer cell growth and survival by depleting NAD+ levels.
References:
Hasmann, M., & Schemainda, I. (2003). FK866, a highly specific noncompetitive inhibitor of nicotinamide phosphoribosyltransferase, represents a novel mechanism for induction of tumor cell apoptosis. Cancer Research, 63(21), 7436-7442.
NAD and Immunotherapy:
Immunotherapy has emerged as a promising approach in cancer treatment. Recent studies have explored the link between NAD and immune system function in the context of cancer. Increasing NAD+ levels in immune cells may enhance their function and improve their ability to recognize and eliminate cancer cells. Although this area of research is relatively new, it holds significant promise for the development of innovative cancer immunotherapies.
References:
Audrito, V., Managò, A., Gaudino, F., Sorci, L., Messana, V. G., Raffaelli, N., … & Deaglio, S. (2019). NAD-Biosynthetic and consuming enzymes as central players of metabolic regulation of innate and adaptive immune responses in cancer. Frontiers in Immunology, 10, 1720. doi:10.3389/fimmu.2019.01720
Conclusion:
The intricate interplay between NAD and cancer cells has unveiled new avenues for cancer research and treatment. From PARP inhibitors to metabolic interventions and immunotherapies, NAD-related strategies are changing the landscape of cancer therapies. However, it’s important to emphasize that ongoing research is necessary to fully understand the potential benefits and complexities of manipulating NAD in the context of cancer treatment. As such, consulting with healthcare professionals and staying informed about the latest scientific developments is crucial for individuals affected by cancer and their treatment options.
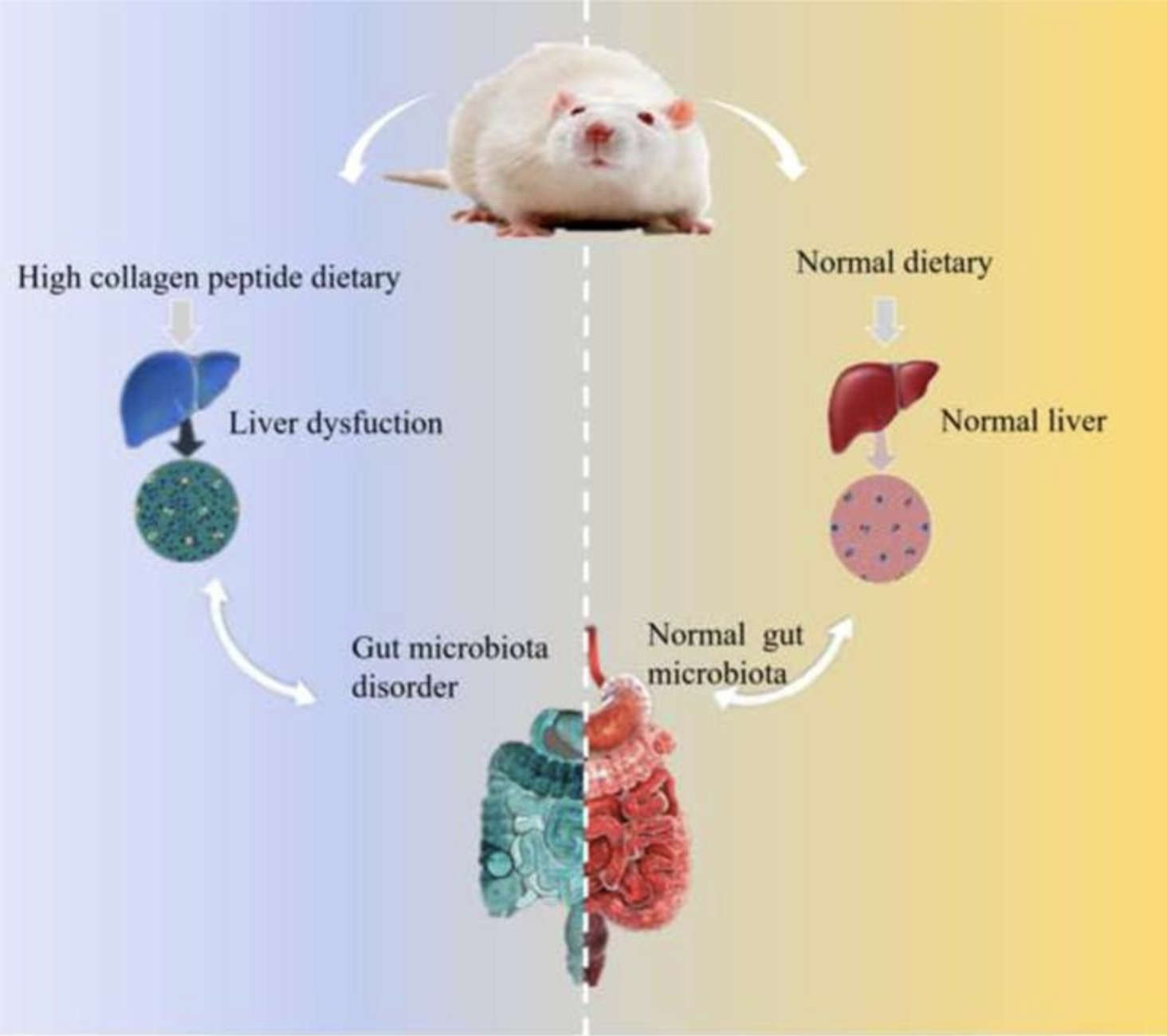
Fengfeng Mei, Zhouwei Duan, Muxue Chen, Jinfeng Lu, Meihui Zhao, Laihao Li, Xuanri Shen, Guanghua Xia, Shengjun Chen, Effect of a high-collagen peptide diet on the gut microbiota and short-chain fatty acid metabolism, Journal of Functional Foods, Volume 75, 2020, 104278, ISSN 1756-4646, https://doi.org/10.1016/j.jff.2020.104278.
Highlights
•A high-dose intake of the collagen peptides increased body weights and caused liver dysfunction.
•A high-dose intake of the collagen peptides could profile the profiles of SCFA metabolism.
•A high-dose intake of the collagen peptides could alter of the gut microbiota community.
Abstract
In this study, a total of 24 male Sprague Dawley rats were randomly divided into 3 group (Collagen peptide of Salmon salar skin group, Ss-SCP; Collagen peptide of Tilapia nilotica skin group, Tn-SCP and Model control group, MC) to investigate the impact of a high-collagen peptide diet on the gut microbiota and host health. After 16 days intervention, the body weights of the Ss-SCP and Tn-SCP intervention groups were significantly increased and the liver index was also remarkably higher than that of the MC group. The acetic acid and propionic acid levels in feces were both significantly increased in the diet high-collagen peptide groups and valerate acid level was lower than that in the MC group. With the intervention of a high-dose collagen peptide diet, the gut microbiota of the groups was shifted with increased abundance of Lactobacillus, Unidentified-Prevotellaceae, Allobaculum, and Parasutterella, whereas the Tn-SCP administration have caused low abundance of Anaerostipes, Blautia, and Fusicatenibacter. The relative abundance of Allobaculum as well as Parasutterella was positively correlated with propionic acid and acetic acid levels, respectively. In addition, Allobaculum abundance was negatively correlated with valerate acid level. The serum valerate acid content was potentially harmful to rat health and significantly increased in the groups intervened with collagen peptide. All together, these results showed that administration of diet high-collagen peptide shifts the gut microbiota in rats and induced a disturbance in short-chain fatty acid metabolism which is potentially harmful to health.
Graphical abstract
Keywords
Collagen peptide
Gut microbiota
SCFA metabolism
Valerate acid
1. Introduction
Collagen peptide as a hydrolysate of collagen has various beneficial effects, such as protecting skin from aging, promoting wound healing (Mei et al., 2020, Zhang et al., 2011b), improving bone health (Song, Zhang, Zhang, & Li, 2019), increasing muscle strength (Hong, Fan, Chalamaiah, & Wu, 2019) reducing obesity (Lee et al., 2017), maintaining blood pressure (O’Keeffe, Norris, Alashi, Aluko, & FitzGerald, 2017), preventing atherosclerosis (Mallavia et al., 2013, McCabe et al., 2013), and modifying lipid metabolism (Blachier et al., 2019, Lin et al., 2012). These positive effects of collagen peptide oral administration on human health have motivated people to consume a high-collagen peptide diet in some areas of the world such as china. Specially, for the people who want to keep skin young and healthy, lose weight or the athletes who want to increase muscle content on a high-protein diet, protein intake is usually two to three times more than the recommended amount (Blachier et al., 2019, Eissa et al., 2019; L. Zhang et al., 2018). However, according to the current recommendation of protein amount recommended by the FAO/WHO at 2007 (FAO/WHO, 2007), the safe complete protein intake level is 0.83 g/kg·bw per day for both men and women, which means that the administration of 49.8 g protein satisfies the daily nutritional requirements of a 60-kg man.
According to the dietary protein consumption beyond the FAO/WHO-recommended amount is increasingly being associated with potentially harmful effects, including kidney diseases (Juraschek, Appel, Anderson, & Miller, 2013), bowel diseases, and hepatic dysfunction (Sun et al., 2019). Low-molecular-weight peptides have better bioavailability and are preferred over the larger peptides and parent proteins (Wang et al., 2019, Wang et al., 2019). In addition, low-molecular-weight collagen peptides, but not the complete proteins, from fish skin possess high immunomodulatory and antioxidant activity and better absorption of collagen than proteins from other sources (Xu, Hong, Wu, & Yan, 2019). Previous studies provided insight into the function of low-molecular-weight collagen peptides from fish skin in assist with clinical problems such as chronic wounds (Yang, Zhang, Li, & Hou, 2018). For better efficacy, 2 g/kg is often chosen as the treatment dose in a murine model (Yang et al., 2018, Zhang et al., 2011b), which is a high-dose collagen peptide intervention. Therefore, we need to better understand the detrimental of a high-dose collagen peptide diet.
Besides the host physiological condition, the gut microbiota also plays a key role in protein metabolism (Blachier et al., 2019, De Sordi et al., 2018, Han et al., 2018, Sun et al., 2019). Protein digestion is a very efficient process that depends on the nature of the protein and the proteins or peptides that undergo catalysis by bacterial proteases and peptidases to release shorter peptides and amino acids (Portune et al., 2016). Amino acid synthesis and metabolism in vivo largely remain to be determined, along with management of the effect of gut microbiota on the body nitrogen metabolism (Neis, Dejong, & Rensen, 2015). The increased transfer of nitrogenous compounds from high dietary protein liable to modify the gut microbiota, metabolic activity, and production of metabolites (Blachier et al., 2019, Fan et al., 2017, Portune et al., 2016). Short chain fatty acids (SCFA), including acetate, propionate, butyrate, and valerate acid, are some of the major metabolites of intestinal microorganisms, which are mainly produced by the fermentation of diet fiber and resistant starch that is not digested and absorbed by the host (Bloemen et al., 2010). Recent studies showed that dietary protein could increase SCFA production via increasing gut bacterial relative abundance, which is associated with SCFA metabolism (Han et al., 2018). Following production, acetate, propionate and butyrate are used for energy metabolism after transportation to the liver (Bloemen et al., 2010). Valerate acid, as a potential deleterious SCFA, is excreted in the feces instead of being absorbed into the blood. The remainder of the SCFAs are oxidized by the liver to prevent systemic, highly toxic concentrations.
Despite several crucial studies investigating the relationship of dietary protein within the recommended amount and the gut microbiota axis(Fan et al., 2017); (Han et al., 2018), the link between overload dietary protein, the gut microbiota, and SCFA metabolism remains unclear, and the potential deleterious effects on a body with excess protein remain unevaluated.
In this study, we administered rats with a high-collagen peptide diet composed of the hydrolysis products from Salmon salar (deep-sea fish, Ss-SCP) and Tilapia nilotica (fresh water fish, Tn-SCP) skin collagen. We analyzed the gut microbiota by 16S rDNA high-throughput sequencing, investigated SCFA metabolism, the liver index, cardiac index, spleen index and altered body weights of rats.
2. Materials and methods
2.1. Preparation of collagen peptides
Salmon salar skin was purchased from the market (Hainan, China). Tilapia nilotica collagen peptide (Tn-SCP) was donated by Hainan SEMNL Biotechnology Co., Ltd. (Hainan, China). All chemicals and reagents used were of analytical grade.
Salmon salar collagen peptide (Ss-SCP) was derived from Salmon salar skin by enzymatic hydrolysis according to the method of Mei et al. (2020). Briefly, for purification, the Salmon salar skin was mixed with 20 times the volume of 0.1 mol/L NaOH for 10 min. Second, at pH 9.5, a complex of 0.5% lipase and protease was added and incubated for 4 h before being mixed with 0.05 mol/L acetic acid pH 4 for 3.5 h at 64 °C. Lastly, the mixture was dried into a powder by spray-drying to Ss-SCP powder (Mei et al., 2020, Zhang et al., 2011a). In addition, Tilapia nilotica collagen peptide (Tn-SCP) was donated by Hainan SEMNL Biotechnology Co., Ltd. (Hainan, China). The collagen peptide samples were purified and the amino acidcomposition as well as the molecular weight distribution was analyzed by the Analysis & Testing Center Jiangnan University (Jiangsu, China).
2.2. Animals and diets
A total of 24 male Sprague Dawley (SD) rats (160±10g) were obtained from TianQin Biotechnology Company of Changsha. Rats were housed in the same conditions, including a constant temperature of 23 ± 1 °C and a 12-hour light–dark cycle (Xia, Yu, et al., 2015). All animals were handled according to the guidelines of the principle of Laboratory Animal Care of China Animal Health and Epidemiology Center (www.cahec.cn).
Before experiments, all rats were accepted a seven day’s acclimation period (Han et al., 2018) with a standard isonitrogeneous full-value chow (Changsha TianQin Biotechnology Co. Ltd., China). After that, SD rats were randomly assigned to three groups (n = 8), i.e. the model control group (MC), a Salmon salar skin collagen peptide (Ss-SCP) administration group, and a Tilapia nilotica skin collagen peptide (Tn-SCP) administration group. Rats in the collagen peptide groups were intragastric administered Ss-SCP and Tn-SCP with the dose of 2 g/kg body weight, while rats in MC were intragastric administered the same dose of physiological saline solution as vehicle. All rats were maintained on standard full-value chow (Changsha TianQin Biotechnology Co. Ltd., China) with 18% protein at the same time and weighted every day. Rats received a medium and long term daily oral administration of collagen peptide until they were sacrificed on day 16. The liver index, cardiac index and spleen index were recorded immediately after sacrificed.
2.3. Collection of blood and fecal samples
On day 16, blood samples were collected after the rats were sacrificed and centrifuged (3000 r/min for 20 min at 4 °C) to separate the serum (Mei et al., 2020). Fresh rat fecal samples were collected with sterile forceps aseptically immediately after defecation. The fresh feces were dropped into liquid nitrogen immediately after collection and stored at −80 °C for microbial diversity and metabolic profiling analysis (Xia, Zhao, et al., 2015).
2.4. Genomic DNA extraction, PCR amplification, high throughput sequencing and sequence data bioinformatics analysis
To investigate the relationship among the diet collagen peptide and gut microbiota, 16S rDNA high-throughput sequencing was used to investigate the change in gut bacteria.
The genomic DNA of the feces was extracted with the Qiagen QIAamp DNA Stool Mini Kit (Qiagen, Germany) according to previous study (Zhang & Zhang, 2018), and then the purity and concentration of DNA were detected by agar-gel electrophoresis. An appropriate amount of sample DNA was taken and diluted to 1 ng/ L with sterile water for subsequent 16S rRNA genes.
The S5-16SV45 hypervariable region of the bacterial 16S rRNA gene was amplified with the 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 907R (5′-CCGT CAATTCCTTTGAGTTT-3′) primers. The PCR products were verified by 2% agarosegel electrophoresis. The amplicon was extracted from 2% agarose gel and purified using QIAquick PCR purification kit (QIAGEN, Hilden, Germany).
16S rDNA amplicon sequencing was used to target the variable regions, and general primers were designed for PCR amplification of the variable region; then, sequencing analysis and species identification were carried out for the hypervariable regions. According to the characteristics of the amplified region, the IonS5TMXL sequencing platform was used to construct a small fragment library for single-end sequencing. To study the species composition of each sample, the Effective Tags of all samples were clustered into Operational Taxonomic Units with 97% consistency (Identity), and the representative sequences of OTUs were annotated with species. The alpha and beta diversity analysis was used to determine the differences among samples (Mei et al., 2020). The UPGMA-tree and Shanon-Weiner index was created using the Qiime analysis software. The raw data of the high-throughput sequencing can be found at the Novogene data platform (https://magic.novogene.com).
Cutadapt (V1.9.1, http://cutadapt.readthedocs.io/en/stable/) was used at first to identify low-quality partly sheared reads, and according to the Barcode these reads were split from the sample data reads. The barcode was removed and the preliminary quality of the reads was assessed to obtain the original data sequence (raw reads). Then, chimeric sequences were identified by searching the annotation database (https://github.com/torognes/vsearch/) for matching chimeric sequences and species, finally the chimeric sequences were removed to obtain the final validated data.
Next, the uparse software was used (Uparse v7.0.1001, http://www.drive5.com/uparse/) to cluster all clean reads with a default of 97% consistency resulting in clustering of the OTUs sequence. At the same time, based on the principles of the algorithm, this program will select the typical sequence of the OTUs. The sequences with the highest frequency were selected as the OTU representative sequences. The Mothur method and SILVA132 SSU rRNA database (http://www.arb-silva.de/) were used on the OTU sequences to conduct the species annotation analysis (setting threshold of 0.8–1), acquire taxonomy information and the classification level community composition of each sample: kingdom (community), phylum, class, order, family, genus, and species. The MUSCLE (Version 3.8.31, http://www.drive5.com/muscle/) software was used for multiple sequence alignment. After that, the predicted functional alterations of the gut microbiota in each group were evaluated using the SILVA SSU Ref NR database. Finally, the data from each sample was homogenized and the least amount of data in the sample was used to standardize the homogenization. The subsequent alpha and beta diversity analysis were all based on the homogenized data. The metagenome data are available at NCBI Sequence Read Archive (SRA) data base and Biosample under accession number PRJNA648676.
2.5. Determination of SCFAs in feces and serum
Serum samples (100 µL) were mixed with 25 µL methanol and 50 µL ethanol for deproteinization. The fecal samples were dispersed in distilled water (170 mg/mL), and mixed for 15 min. The supernatant of the mixture was collected after centrifugation (3000 r/min for 15 min at 4 °C) to further analyze the SCFAs. SCFAs (acetate, propionate, butyrate and Valerate acid in feces and serum) were studied with the Agilent 7890A gas chromatograph (Agilent, C) equipped with a flame ionization detector and a HP-FFAP column (30 mm × 0.32 mm × 0.25 µm, Agilent, USA) to evaluate the SCFA content of samples. The standard acetic acid, propionic acid, and Valerate acid (>99%) for gas chromatograph analysis were purchased from Aladdin Bio-Chem Technology Co., Ltd., China (Shanghai, China). All samples were prepared for SCFA analysis according to the following program: an initial temperature of 80 °C for 0.5 min, 80–150 °C at 4 °C/min, and finally 150–230 °C at 20 °C/min for 10 min. The detector temperature was 250 °C.
2.6. Statistical analysis
Data are presented as the mean ± standard error of the mean. The sequence data bioinformatics analysis was used Source Tracker analyses and micro PITA analyses. T-test and Wilcox rank sum test were performed for 2 groups. R software (Version 2.15.3) was used to draw PCA, PCoA, and NMDS diagrams. Other distribution and statistical comparison analyses of each group were performed using an ANOVA test in the Statistical Product and Service Solutions software. The analysis probability values of P < 0.05 were accepted as statistically significant.
3. Results
3.1. Composition and characteristics of collagen peptides
Our previous study has found that the average molecular weight distribution of Tn-SCP was 460 Da and the average molecular weight of Ss-SCP was consistent with Tn-SCP (Mei et al., 2020).
As presented in Table 1, the amino acid composition of Tn-SCP and Ss-SCP were different. The Tn-SCP was rich in Gly > Glu > Pro > Ala > Hyp > Arg > Asp and the Ss-SCP was rich in Gly > Glu > Pro > Hyp > Asp > Ala > Arg. In addition, the essential amino acids (EAA) of Tn-SCP and Ss-SCP accounted for 15.81% and 22.20% of the total amino acids (EAA/TAA), respectively. The ratio of essential amino acids to non-essential amino acids (EAA/NEAA) of Tn-SCP and Ss-SCP were 18.77% and 28.55%, respectively. These results indicated that the Tn-SCP and Ss-SCP conformed to the characteristics of collagen peptides. However, Ss-SCP and Tn-SCP were composed of incomplete proteins according to the FAO/WHO, which defines incomplete proteins as EAA/TAA ≥ 0.4 and high-quality proteins as EAA/NEAA ≥ 0.6.
3.2. Diet high-dose of collagen peptide directly influenced host health
To investigate the impact of the high-collagen peptide diet on rats, the general information of each group of rats were recorded. During the days of feeding, the daily food intake per rat was slightly different. Approximately, per rat in each group, 7.2 g crude protein from full-value chow was consumed (Table 2).
The general full-value chow daily intake information was recorded to investigate the average daily basic nutrient intake.
The average initial weight of all rats was 160 g ±10 g (Fig. 1A) and the final body weight of rats in the Tn-SCP and Ss-SCP groups was significantly higher than the MC group (Fig. 1A-B) (P < 0.05). In addition, the viscera index of the cardiac, liver (Sun et al., 2019) and spleen, which is directly correlated with amino acid metabolism (Blachier et al., 2019, Juraschek et al., 2013), was recorded immediately after sacrifice. The cardiac index was not significantly different between each group (Fig. 1C) (P > 0.05). In addition, compared with the MC, the spleen index in the Tn-SCP group was significantly increased but not significantly different from the Ss-SCP group, which might be associated with the amino acid composition of the peptides. The liver weight in the collagen peptide groups was significantly higher than in the MC (Fig. 1D) (Ss-SCP vs MC, P < 0.01; Tn-SCP vs MC, P < 0.001). Histologically, the liver histological image showed that compared with the MC group the arrangement of hepatocytes in the collagen group was relatively loose, and large number of cells were swollen near the portal area, which showed obvious hepatocyte injury. Thus, these data provide compelling evidence that intervention with a high collagen peptide diet directly influenced host health and might induce weight gain of the host.
3.3. Diet high-dose collagen peptide induced a credible change of gut microbiota transfer
After 16S rDNA high-throughput sequencing, an average of 85,231 reads were detected per sample, and 80,095 effective reads were obtained through quality control on average, with the quality control effective rate reaching 94.04%. Sequence clustering was conducted to transform the read data into OTUs with 97% consistency, and a total of 612 OTUs were obtained (Table 3).
Table 3. Information of 16S rDNA high-throughput sequencing.
Empty Cell
Raw reads
Clean reads
Base (nt)
Avglen (nt)
Q20
GC%
Effective (%)
MC
84,643
80,099
29,865
372
84
53
94.68
Ss-SCP
85,731
80,070
298,265
372
82
53
93.48
Tn-SCP
85,317
80,115
2,961,458
369
77
53
99.95
PS
85,231
80,095
29,688,473
370
75.83
53.29
94.04
Data preprocessing and quality control. The OTU levels of the large intestine reactor are different. Samples were collected from the rats immediately after sacrifice (n = 21). Raw reads were filtered for sequences of low-quality. Clean reads refer to the sequence that is finally used for subsequent analysis after filtering chimeras. Base refers to the Base number of final Clean reads. AvgLen refers to the average length of the Clean reads. Q20 refers to the percentage of bases whose base mass value in Clean reads is greater than 20 (sequencing error rate is less than 1%). GC (%) represents GC base content in the Clean reads. Effective (%) represents the percentage of the number of Clean reads versus the number of Raw reads. MC, model control group. Ss-SCP, Salmon salar skin collagen peptide group. Tn-SCP, Tilapia nilotica skin collagen peptide group. PS, Average value per sample in each group.
Rarefaction curves (Fig. 2A) and rank abundance (Fig. 2B) showed that the sample size and sequencing depth were increased and no additional OTUs were detected indicating that the gut bacteria sequencing results, which reflect sample diversity and richness of the microbiota, were credible.
Principal coordinate analysis (PCoA) (Fig. 2C) revealed that the principal components of PC1 and PC2 contributed about 67.54% and 13.35%, respectively. The PCoA indicated that there was a slight difference between the MC and Ss-SCP groups, however, the PC1 and PC2 in the Tn-SCP group were significantly different in comparison to the MC. The Tn-SCP group was mainly concentrated in the third and fourth quadrants, while the Ss-SCP and MC groups were mainly concentrated in the first and second quadrants. Nonmetric multidimensional (Fig. 2D) analysis further demonstrated that there was a total separation between the gut microbiota profiles of the MC and Tn-SCP groups, but there was similarity between the MC and Ss-SCP groups. Observed_species index and Shannon index (Fig. 2E–F) was significantly different between Mc VS Tn-SCP (P < 0.05) and showed no statistical difference between MC VS Ss-SCP and Ss-SCP VS Tn-SCP (P > 0.05). Collectively, the overall results indicated that the gut microbiota sequencing results in the groups with the high-collagen peptide diet, which reflect sample diversity and richness of microbiota, were credible and the extent of the gut microbiota transformation might be related to the amino acid composition of the collagen peptides.
3.4. Composition of gut microbiota
The gut microbiota at different classification levels under a high-collagen peptide diet was profiled using 16S rDNA high-throughput sequencing. Bacterial DNA profiles (obtained using the weighted Unifrac method with an arithmetic mean dendrogram) displayed a clear segregation between the Tn-SCP and MC and a slight difference between the Ss-SCP and MC (Fig. 3A). The relative abundance of the gut microbiota at the phylum level showed a significant alteration of microbiota in the MC versus Tn-SCP, including Firmicutes (61% in MC vs. 39% in Tn-SCP) (P < 0.01), Bacteroidetes (37% in MC vs. 58.7% in Tn-SCP) (P < 0.01) and Proteobacteria (1.34% in MC vs. 1.12% in Tn-SCP) (P < 0.05), but was insignificant between the MC and Ss-SCP (Fig. 3B–C). At the genus level, the gut microbiota showed a striking difference in those three groups, especially in Lactobacillus, Unidentified-Prevotellaceae, Blautia, Fusicatenibacter, Allobaculum and Anaerostipes (Fig. 3D, Table 4). Despite the fact that there were six bacteria altered in the collagen peptide group compared with the MC, only Allobaculum, Anaerostipes and Fusicatenibacter were significantly changed in both the Tn-SCP and Ss-SCP groups (Table 4) (P < 0.01). Interestingly, the Lactobacillus and Unidentified-Prevotellaceae were sharply changed in the Tn-SCP group but only had a slight difference in the Ss-SCP group compared to the MC (Table 4). All in all, these results indicated that the gut microbiota changes were significantly associated with the composition and characteristics of the collagen peptides.
Table 4. Relative abundance at the genus level of top nine bacteria.
Empty Cell
MC
Ss-SCP
Tn-SCP
Lactobacillus
0.281307 ± 0.182548
0.2478 ± 0.129965
0.132685 ± 0.114587*
Unidentified-Prevotellaceae
0.151382 ± 0.144975
0.103146 ± 0.078058
0.253448 ± 0.096791*
Blautia
0.092052 ± 0.062671
0.109049 ± 0.100574
0.035863 ± 0.032427*
Fusicatenibacter
0.043249 ± 0.039342
0.027526 ± 0.019437*
0.018091 ± 0.019437**
Alloprevotella
0.055212 ± 0.034787
0.057975 ± 0.041895
0.052572 ± 0.030562
Allobaculum
0.010166 ± 0.010426
0.03051 ± 0.036844*
0.036738 ± 0.02502*
Anaerostipes
0.01548 ± 0.033049
0.002695 ± 0.002277*
0.003661 ± 0.008809*
Phascolarctobacterium
0.048534 ± 0.017869
0.052403 ± 0.026639
0.049421 ± 0.023741
Bacteroides
0.021055 ± 0.0085
0.019016 ± 0.015015
0.031242 ± 0.018069
Data are the mean ± standard error of the mean. *P < 0.05, **P < 0.01, compared with the MC group.
3.5. The altered gut microbiota was linked to the concentration of SCFAs
To investigate the impact of a high-collagen peptide diet on the gut microbiota and a direct link with metabolism, we further analyzed the SCFA content and screened out the most remarkable data for analysis. In fecal samples, acetate was the most abundant SCFAs followed by propionate, and the concentration of acetate (Fig. 4B) or propionate (Fig. 4D) were higher in the collagen peptide intervention groups than in the MC group (acetate, P < 0.001 for Ss-SCP and Tn-SCP; propionate, P < 0.05 for Ss-SCP, P < 0.01 for Tn-SCP). The butyrate concentration (Fig. 4E) in Ss-SCP were significantly higher than MC group (P < 0.01) but there was no statistic difference between MC and Tn-SCP groups. It is worth noting that the valerate acid levels in the Tn-SCP and Ss-SCP groups were both significantly decreased in comparison with that in the MC group (Fig. 4C) (P < 0.05). We next detected valerate acid in serum; however, the valerate acid levels in the collagen peptide intervention groups were higher than those in the MC group (Fig. 4F) (P < 0.01 for Ss-SCP, P < 0.001 for Tn-SCP). To illustrate the change in the gut microbiota, a correlation heatmap is shown in Fig. 4A and Fig. S1. As shown in Fig. 4, at the genus level, the significantly altered bacteria were Allobaculum, Faecalibacterium, unidentified-Prevotellaceae, Parabacteroides, Sellimonas, Lachnoclostridium, Roseburia, unidentified-Lachnospiraceae and Parasutterella (relative abundance Ss-SCP > Tn-SCP > MC, P < 0.01 for Tn-SCP and Ss-SC in comparison to MC). We next addressed the potential contribution of the altered gut microbiota to the variety of SCFAs in rats fed the high-collagen peptide diet (Table 5). Using Pearson’s correlation, we analyzed the correlation between the concentrations of the significantly changed SCFAs and the relative abundance of the main gut microbiota at the genus level. Allobaculum, which was highly abundant in both the Tn-SCP and Ss-SCP groups (Table 4), was significantly positively correlated with propionic acid level (correlation coefficient was 0.953, P < 0.05) and butyrate acid level (correlation coefficient was 0.835, P < 0.05) (Table 5)and negatively correlated with valerate acid level (correlation coefficient was −0.977) (Table 5) (P < 0.05). In addition, Parasutterella, with a high abundance in the collagen peptide intervention groups was positively correlated with acetate level (correlation coefficient was 0.954).
Table 5. Pearson’s correlation coefficient between the relative abundance of bacteria and levels of SCFAs.
Bacteria Cenera
Pearson’s correlation coefficient
Acetate
Propionic
Valerate
Butyrate
Allobaculum
0.936065
0.953*
−0.977*
0.835*
Parasutterella
0.954*
0.56377
−0.90137
0.648
Faecalibacterium
−0.26846
0.385874
0.127487
−0.41
unidentified_Prevotellaceae
0.070324
0.673053
−0.2127
−0.239
Parabacteroides
0.530212
0.941413
−0.64636
0.17
Sellimonas
0.411707
−0.24122
−0.27671
0.33
Lachnoclostridium
−0.09859
−0.69376
0.240331
0.229
Roseburia
−0.26642
−0.80615
0.40194
−0.122
unidentified_Lachnospiraceae
0.082919
−0.55214
0.060904
−0.259
*P < 0.05, **P < 0.01, compared with the MC group.
3.6. Functional prediction changes in the gut microbiota
The predicted microbial functions including genetic-information processing, human diseases, metabolism, organismal systems, cellular processes and environmental information processing were analyzed in the present study (Fig. 5A). The functional pathways of the Ss-SCP group were similar to the MC group and both of those groups showed a high abundance in the cellular process pathway and environmental information processing pathways. The gut microbiota functions in the Tn-SCP group were totally different than the other groups. The Tn-SCP group showed increases in genetic information processing, human diseases, metabolism and organismal system metabolic pathways and decreases in the cellular processes and environmental information processing pathway compared with the MC group (P < 0.05). Collectively, the overall gut microbiota predicted function indicated whether the gut microbiota function would be altered and may be related to the composition of collagen peptide or the molecular weight.
4. Discussion
The investigation of the impacts on gut health with an intervention of protein diets has shown a surge of interest. Previous studies were focused on the benefits of a dietary protein within an appropriate dose (Fan et al., 2017, Han et al., 2018, Zhu et al., 2015), especially the modulation of the gut microbiota and production of SCFAs. The role of high content of protein in the diet and how that would modulate the gut microbiota and levels of SCFAs remains unclear.
In this study, the interaction of a high-collagen peptide diet with the gut microbiota and SCFA metabolism were investigated by giving rats a diet with high Tn-SCP and Ss-SCP collagen peptides and then evaluating the influence on the body’s metabolism and gut bacteria diversity. From a nutritional perspective, the Tn-SCP and Ss-SCP collagen peptides were not complete proteins according the FAO/WHO, and the composition of the Ss-SCP were closer to a complete protein composition than the Tn-SCP. The average molecular weight of the Tn-SCP and Ss-SCP were less than 500 Da, indicating that both peptides were low molecular weight and would facilitate digestion and absorption. During the experiment, the rats were all healthy and had no clinical symptoms such as diarrhea. On average, the rats of each group took in equal amounts of chow, but the rats in the Tn-SCP and Ss-SCP group gained significantly more weight compared to the MC. However, the collagen peptide diet with an advisable dose has been shown to have positive effects on inhibition of preadipocyte differentiation and has ameliorated obesity in rats with a high fat diet (Lee et al., 2017). In vivo, proteins can be converted to lipids via amino acid metabolism; the liver (Li et al., 2018) and spleen (Gong et al., 2014) are the main sites of lipid anabolism. Interestingly, the spleen index showed that the spleen weight of rats in the Tn-SCP group were higher than in the MC (P < 0.01), but the spleen index in the Ss-SCP group, which had a closer amino acid composition with complete protein, was only slightly different compared to the MC (P > 0.05). These results confirmed that the amino acid composition of the collagen peptides might have a direct link with the shifted spleen lipid metabolism. The liver index was significantly higher in the diet high-collagen peptide intervention groups (P < 0.001 for Tn-SCP, P < 0.01 for Ss-SCP) than in the MC group and the H&E staining showed that the arrangement of hepatocytes in the collagen group was relatively loose, which agrees with recently study from Islam et al (Islam, Ravandi, & Aukema, 2018) that a high-protein diet is associated with a physiologically enlarged liver. Thus, in the high-collagen peptide intervention condition, excessive amino acid metabolism in the liver can lead to liver enlargement, which can lead to liver dysfunction.
We speculated that the liver function may be directly linked to the exchange of SCFAs. Previous studies reported the correlation between liver function and SCFA metabolism (Bloemen et al., 2009, Bloemen et al., 2010) SCFAs are the major metabolites produced by gut microbiota and are key signaling molecules in various biochemical reaction processes (Koh, De Vadder, Kovatcheva-Datchary, & Backhed, 2016). SCFAs are absorbed and metabolized by the colonic epithelium, peripheral muscle tissue, and liver following their production (Blachier et al., 2019, Bloemen et al., 2010). SCFAs enter the portal circulation of the liver, directly regulating various pathways involved in the metabolism of fatty acids and cholesterol. In our present study, compared with MC group, the butyrate level of feces in Ss-SCP was significantly higher but in Tn-SCP was no statistical difference. Butyrate as an energy source is largely utilized by the colonic epithelium (Blachier et al., 2019, Han et al., 2018, Lin et al., 2012). Previous study showed that the butyrate could suppress diet-induced obesity in Free fatty acidreceptor gene lacked mice(Lin et al., 2012) and regulated the energy homeostasis(Byrne, Chambers, Morrison, & Frost, 2015), which indicated that the butyrate is a beneficial SCFAs. As the Ss-SCP were closer to a complete protein composition, we suspect that composition of consumed protein is tightly relative with the butyrate produce. In addition, significantly high acetate and propionate levels, but low valerate acid levels in the feces of the Tn-SCP and Ss-SCP intervention groups were observed.
In this experiment, due to the low content of acetic acid and propionic acid in the serum, we did not detect acetate and propionic acid under the existing experimental conditions, but we observed a significant difference in valerate acid levels in contrast to that in the serum propionate levels in the diet collagen peptide groups compared to the MC group. Acetate and propionate, as the beneficial acids, may be in a dynamic equilibrium due to their hilar circulation, oxidization, and use as an energy source in vivo. The remaining propionate is used in gluconeogenesis and inhibits the synthesis of cholesterol, whereas acetate is used to synthesize the long-chain fatty acids, glutamine and beta hydroxybutyrate (Bloemen et al., 2009, Bloemen et al., 2010). The absorption mechanism of valerate acid in vivo remains unclear, as the high systemic concentration of valerate acid is toxic and may exert an adverse influence on the patient. Previous studies observed that propionate and acetate play an important role in appetite and energy regulation (Byrne et al., 2015), by regulating the free fatty acid receptor 3 (Lin et al., 2012) and reducing appetite via a homeostatic mechanism (Byrne et al., 2015). Thus, we hypothesize that the increased body weight gain in the collagen peptide intervention groups with significantly high levels of propionate and acetate were caused by liver dysfunction related to the high systemic concentration of valerate acid, which is worthy of further investigation.
The profiles of the SCFAs are tightly correlated with the gut microbiota diversity (Fan et al., 2017, Guardia-Escote et al., 2019, Wu et al., 2019). Despite the SCFAs showing a uniform change in the diet high-collagen peptide groups, the UPGMA analysis displayed a clear segregation between the Tn-SCP group and the MC and a slight difference between the Ss-SCP group and the MC. These data suggested that the intervention diet containing Tn-SCP strongly altered the structure of the gut bacterial communities while the Ss-SCP group may have a composition of gut microbiota similar to that of the MC group at the phylum level. Regarding the different results observed in the collagen peptide groups, the composition of amino acids might have greatly contributed to the change in the gut microbiota. The extent of the change in the gut microbiota would be less in the Ss-SCP group, which had an amino acid composition similar to that of the complete protein, according to the FAO/WHO.
For the major bacteria, Allobaculum, a member of the phyla Firmicutes, was highly abundant in both the Tn-SCP and Ss-SCP intervention groups and has been confirmed as a bacteria linked to the butyrate content in vivo (Han et al., 2018, McLeod et al., 2019). In the present study, the abundance of Allobaculum presented a remarkable positive correlation with the levels of propionic acid (P < 0.05) and butyrate acid (P < 0.05) and a negative correlation with the levels of valerate acid (P < 0.05). Parasutterella, which is presented in the heatmap graph as the most differentially abundant bacteria in the high-collagen peptide groups was also significantly positively correlated with the production of acetic acid. To our knowledge, previous studies have suggested that Parasutterella is a potential harmful bacteria (Wan et al., 2019), but little is known about its role in SCFAs metabolism. Thus, we hypothesized that the levels of SCFAs in the collagen peptide groups could be ascribed to the shifted Parasutterella and Allobaculumbacterial metabolism caused by the interaction with the high-collagen peptide diet.
Several previous studies have shown that the Allobaculum are potentially beneficial to the host and are found to have richer abundance in fat rats with a high fat diet (Bai et al., 2018, Everard et al., 2014) or rats with a high food intake (Han et al., 2018). However, Allobaculum were in high abundance in the collagen peptide groups with an overweight tendency. The predicted function of the gut microbiota in the Tn-SCP group was totally different than the MC group, and the Ss-SCP group also increased the unclassified pathway in comparison to the MC group. Furthermore, we hypothesized that the body weight result might be linked with the interaction of Allobaculum and other gut bacteria. For the gut bacteria profile, it was notable that the Tn-SCP and Ss-SCP groups both showed a significant abundance of Fusicatenibacter and Anaerostipes compared to the MC group, while only the Tn-SCP intervention group presented a lower abundance of Lactobacillus and Blautia. Fusicatenibacter and Anaerostipes, as the neutral bacteria, were increased via the intervention of the collagen peptide. Lachnospiraceae, which belongs to the Firmicutes, is a typical inflammatory bacterium that exists in the intestinal tract. In the obese mouse model, the relative abundance of Lachnospiraceae is increased with early subtherapeutic antibiotics, and is associated with metabolic shifts including diabetes (Kameyama & Itoh, 2014), obesity (Meehan & Beiko, 2014), and colitis (Zeng, Ishaq, Zhao, & Wright, 2016). Abundantstudies indicate that the Blautia are considered a health-promoting bacteria, which are associated with obesity and improved abundance in the high fat diet-induced obese rats (Bai et al., 2018). Furthermore, we speculate the altered abundance of Lachnospiraceae and Blautia correlated with the changes in the functional pathways altered in the Tn-SCP group in comparison with the MC and Ss-SCP groups including genetic information processing, human diseases, metabolism and organismal system metabolic pathways and decreases in the cellular processes and environmental information processing.
Our main finding was that that high-collagen peptide diet had a negative effect on the gut microbiota and SCFA metabolism. Briefly, the intestinal microbiota, as well as the SCFA metabolism of rats with excessive peptide, were shifted, and increased the risk of obesity to some extent.
5. Conclusion
In conclusion, these studies demonstrated that a high intake of the collagen peptides Tn-SCP and Ss-SCP could shift the profiles of SCFA metabolism via altering the gut microbiota. The amino acid composition tightly relative with the profiles of gut microbiota and SCFAs. In addition, the physical condition of the rats revealed that the high-collagen peptide diet induced rat weight gain and might cause liver dysfunction. This suggests that the daily peptide intake should be within the national limit and choose the complete protein. The long-term excessive consumption of peptide can cause dysfunction of metabolism and the intestinal flora.
6. Ethics statements
The study was performed in accordance with the Experimental Animal Administration regulations issued by the State Committee of Science and Technology of the People’s Republic of China and the Guidelines for Care and Use of Laboratory Animals of Hainan Medical College. The ethical approval reference number of the study is HYXK 20190035. All the procedures for the care of the rats were in accordance with the institutional guidelines for animal use in research.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by Fund of Key Laboratory of Aquatic Product Processing, Ministry of Agriculture and Rural Affairs, China (no. NYJG202001), China Agriculture Research System (no. CARS-46), Special Scientific Research Funds for Central Non-profit Institutes, Chinese Academy of Fishery Sciences (no. 2020TD73), Marine and Fisheries Bureau of Haikou (no. HHCL201804), Innovative Research Projects of Graduate Students of Hainan Province (no. hys2019-73) and the Scientific Research Foundation of Hainan University (no. kyqd1662).
Appendix A. Supplementary material
The following are the Supplementary data to this article:
Blachier et al., 2019F. Blachier, M. Beaumont, K.J. Portune, N. Steuer, A. Lan, M. Audebert, …, Y. SanzHigh-protein diets for weight management: Interactions with the intestinal microbiota and consequences for gut health. A position paper by the my new gut study groupClinical Nutrition, 38 (3) (2019), pp. 1012-1022, 10.1016/j.clnu.2018.09.016View PDFView articleView in ScopusGoogle Scholar
Byrne et al., 2015C.S. Byrne, E.S. Chambers, D.J. Morrison, G. FrostThe role of short chain fatty acids in appetite regulation and energy homeostasisInternational Journal of Obesity (London), 39 (9) (2015), pp. 1331-1338, 10.1038/ijo.2015.84 View PDF This article is free to access.View in ScopusGoogle Scholar
Everard et al., 2014A.L.V. Everard, N. Gaïa, M. Johansson, M. Ståhlman, F. Backhed, N.M. Delzenne, J. Schrenzel, P. François, P.D. CaniMicrobiome of prebiotic-treated mice reveals novel targets involved in host response during obesityIsme Journal (2014), 10.1038/ismej.2014.45 View PDF This article is free to access.Google Scholar
Fan et al., 2017P. Fan, P. Liu, P. Song, X. Chen, X. MaModerate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig modelScientific Reports, 7 (2017), p. 43412, 10.1038/srep43412 View PDF This article is free to access.View in ScopusGoogle Scholar
Guardia-Escote et al., 2019L. Guardia-Escote, P. Basaure, J. Biosca-Brull, M. Cabré, J. Blanco, C. Pérez-Fernández, …, M.T. ColominaAPOE genotype and postnatal chlorpyrifos exposure modulate gut microbiota and cerebral short-chain fatty acids in preweaning miceFood and Chemical Toxicology, 110872 (2019), 10.1016/j.fct.2019.110872View article Google Scholar
Li et al., 2018D. Li, L. Li, T. Xu, T. Wang, J. Ren, X. Liu, Y. LiEffect of low molecular weight oligopeptides isolated from sea cucumber on diabetic wound healing in db/db miceMarine Drugs, 16 (1) (2018), 10.3390/md16010016View article Google Scholar
Lin et al., 2012H.V. Lin, A. Frassetto, E.J. Kowalik Jr., A.R. Nawrocki, M.M. Lu, J.R. Kosinski, …, D.J. MarshButyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanismsPLoS ONE, 7 (4) (2012), Article e35240, 10.1371/journal.pone.0035240View article View in ScopusGoogle Scholar
McLeod et al., 2019K.H. McLeod, J.L. Richards, Y.A. Yap, E. MariñoChapter 7 – Dietary short chain fatty acids: How the gut microbiota fight against autoimmune and inflammatory diseasesR.R. Watson, V.R. Preedy (Eds.), Bioactive food as dietary interventions for arthritis and related inflammatory diseases (2nd ed.), Academic Press (2019), pp. 139-159View PDFView articleGoogle Scholar
Mei et al., 2020F. Mei, J. Liu, J. Wu, Z. Duan, M. Chen, K. Meng, …, M. ZhaoCollagen peptides isolated from salmo salar and tilapia nilotica skin accelerate wound healing by altering cutaneous microbiome colonization via upregulated NOD2 and BD14Journal of Agriculture and Food Chemistry, 68 (6) (2020), pp. 1621-1633, 10.1021/acs.jafc.9b08002View article View in ScopusGoogle Scholar
Wang et al., 2019G. Wang, N. Xu, L. Yang, F. Zheng, L. Sai, J. Zhou, S. YangCommunity acquired Stenotrophomonas maltophilia discitis: Diagnosis aided by shotgun metagenomic sequencingInternational Journal of Infectious Diseases, 81 (2019), pp. 1-3, 10.1016/j.ijid.2019.01.032View PDFView articleGoogle Scholar
Wang et al., 2019K. Wang, M. Liao, N. Zhou, L. Bao, K. Ma, Z. Zheng, …, H. LiuParabacteroides distasonis alleviates obesity and metabolic dysfunctions via production of succinate and secondary bile acidsCell Reports, 26 (1) (2019), pp. 222-235 e225, 10.1016/j.celrep.2018.12.028View articleGoogle Scholar
Wu et al., 2019R. Wu, X. Tang, X. Kang, Y. Luo, L. Wang, J. Li, …, D. LiuEffect of a Chinese medical nutrition therapy diet on gut microbiota and short chain fatty acids in the simulator of the human intestinal microbial ecosystem (SHIME)Journal of Functional Foods, 62 (2019), Article 103555, 10.1016/j.jff.2019.103555View PDFView articleView in ScopusGoogle Scholar
Xia et al., 2015G. Xia, Z. Yu, Y. Zhao, Y. Wang, S. Wang, M. He, …, C. XueSialoglycoproteins isolated from the eggs of Carassius auratus prevents osteoporosis by suppressing the activation of osteoclastogenesis related NF-κB and MAPK pathwaysJournal of Functional Foods, 17 (2015), pp. 491-503, 10.1016/j.jff.2015.05.036View PDFView articleView in ScopusGoogle Scholar
Xia et al., 2015G. Xia, Y. Zhao, Z. Yu, Y. Tian, Y. Wang, S. Wang, …, C. XuePhosphorylated peptides from antarctic krill (Euphausia superba) prevent estrogen deficiency induced osteoporosis by inhibiting bone resorption in ovariectomized ratsJournal of Agriculture and Food Chemistry, 63 (43) (2015), pp. 9550-9557, 10.1021/acs.jafc.5b04263View article View in ScopusGoogle Scholar
Zhang et al., 2011aZ. Zhang, J. Wang, Y. Ding, X. Dai, Y. LiOral administration of marine collagen peptides from Chum Salmon skin enhances cutaneous wound healing and angiogenesis in ratsJournal of the Science of Food and Agriculture, 91 (2011), p. 2173View in ScopusGoogle Scholar
Zhang et al., 2011bZ. Zhang, J. Wang, Y. Ding, X. Dai, Y. LiOral administration of marine collagen peptides from Chum Salmon skin enhances cutaneous wound healing and angiogenesis in ratsJournal of the Science of Food and Agriculture, 91 (12) (2011), pp. 2173-2179, 10.1002/jsfa.4435View article View in ScopusGoogle Scholar
Zhu et al., 2015Y. Zhu, X. Lin, F. Zhao, X. Shi, H. Li, Y. Li, …, G. ZhouErratum: Meat, dairy and plant proteins alter bacterial composition of rat gut bacteriaScientific Reports, 5 (2015), p. 16546, 10.1038/srep16546 View PDF This article is free to access.View in ScopusGoogle Scholar
Xu, S., Xu, H., Wang, W. et al. The role of collagen in cancer: from bench to bedside. J Transl Med17, 309 (2019). https://doi.org/10.1186/s12967-019-2058-1
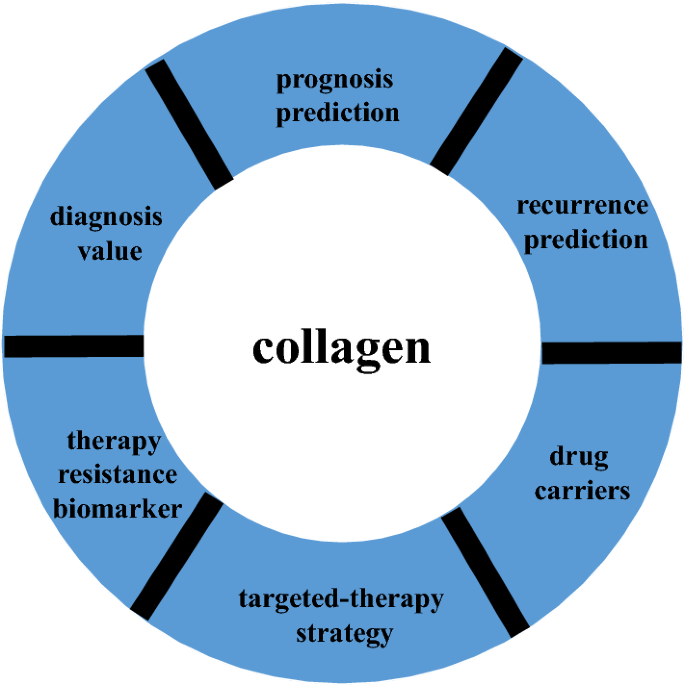
Collagen is the major component of the tumor microenvironment and participates in cancer fibrosis. Collagen biosynthesis can be regulated by cancer cells through mutated genes, transcription factors, signaling pathways and receptors; furthermore, collagen can influence tumor cell behavior through integrins, discoidin domain receptors, tyrosine kinase receptors, and some signaling pathways. Exosomes and microRNAs are closely associated with collagen in cancer. Hypoxia, which is common in collagen-rich conditions, intensifies cancer progression, and other substances in the extracellular matrix, such as fibronectin, hyaluronic acid, laminin, and matrix metalloproteinases, interact with collagen to influence cancer cell activity. Macrophages, lymphocytes, and fibroblasts play a role with collagen in cancer immunity and progression. Microscopic changes in collagen content within cancer cells and matrix cells and in other molecules ultimately contribute to the mutual feedback loop that influences prognosis, recurrence, and resistance in cancer. Nanoparticles, nanoplatforms, and nanoenzymes exhibit the expected gratifying properties. The pathophysiological functions of collagen in diverse cancers illustrate the dual roles of collagen and provide promising therapeutic options that can be readily translated from bench to bedside. The emerging understanding of the structural properties and functions of collagen in cancer will guide the development of new strategies for anticancer therapy.
Background
Cancer continues to receive increasing attention from the academic community because it was the third most common cause of death worldwide in 2018. A total of 18.1 million new cancer cases and 9.6 million cancer deaths were evaluated in 2018 [1], and there are predicted to be 1,762,450 additional cancer cases and 606,880 cancer deaths in the United States in 2019 [2]. Despite various cancer-related guidelines for diagnosis, treatment, and follow-up, improving the long-term prognoses of certain cancer patients remains difficult. Cancer treatment strategies with highly effective response rates still need to be explored. An increasing amount of recent research has concentrated on the function of the tumor microenvironment in favoring cancer progression. In addition, cancer cells exhibit multiple hallmarks of cancer progression, including the recruitment of various cells to form a tumor microenvironment [3], which consists of varying functional stromal cell subtypes and matrix protein polymers [4]. The most abundant matrix protein polymers are collagens, which increase tumor tissue stiffness, regulate tumor immunity, and promote metastasis [5, 6]. In addition, extensive collagen deposition is the main pathological characteristic of some cancers, for which sufficient therapeutic applications are lacking, resulting in the poor survival outcomes of patients [7]. Herein, we summarize the current understanding of the key basic and clinical functions of collagen in cancer and provide clues regarding promising treatments for modifying the tumor matrix.
Physiological and physicochemical properties of collagen
Collagen is a type of right-handed helix glycoprotein that contains three homologous or nonhomologous left-handed helix α chains. These α chain amino acid sequences are characterized by glycine–X–Y repeats with or without interruptions, with X and Y most likely being proline or hydroxyproline, and the hydroxyproline content of collagen contributes to its thermal stability [8].
Nascent α chains by different genes are encoded first to compose the N-terminus. The next step of assembly into a three-helix structure begins with the C-terminus of the nascent α chains to form procollagen, which is accompanied by certain chaperone proteins including heat shock protein 47, prolyl-hydroxylase, and protein disulfide isomerase to ensure precise alignment [9]. Hydroxylation and glycosylation in the endoplasmic reticulum are two main modifications that occur after translation, and the hydroxylation modification is regulated by vitamin C and pyruvate metabolism [10, 11]. Then, procollagen is hydrolyzed to form collagen by procollagen N-proteinase and C-proteinase within Ca2+ surrounding the endoplasmic reticulum along with the chaperone heat shock protein 47 and protein disulfide isomerase. This important hydrolysis reaction is the rate-limiting step of collagen biosynthesis. In addition, endopeptidases and metalloproteinases can also excise procollagen at both the N-terminus and C-terminus, and the removed propeptides can conversely regulate the amount of procollagen, further influencing collagen production [12, 13].
Collagen is released into the extracellular matrix (ECM) to form a fibril supramolecular assembly that may start in Golgi-to-membrane carriers after procollagen excision or be localized at the plasma membrane of fibroblasts. The stability of collagen assembly is influenced by intramolecular and intermolecular linkages, particularly covalent linkages, chiefly including lysyl oxidase (LOX) crosslinks [14], glycosylation crosslinks [15], and transglutaminase crosslinks [16], which vary across collagen types.
Different collagens in the ECM are finally degraded by various matrix metalloproteinases (MMPs) belonging to the zinc-dependent endopeptidase family, by proline oxidase, or by sheddases that release the soluble ectodomain of membrane collagens [17, 18].
In general, the 28 known collagen types are classified into four subfamilies on the basis of their supramolecular assemblies, including fibril-forming collagens (I, II, III, V, XI, XXVI, XXVII); fibril-associated collagens with interrupted triple helices (FACITs: IX, XII, XIV, XVI, XIX, XX, XXI, XXII, XXIV), which characteristically link to the surface of collagen fibrils rather than form fibrils by themselves; network-forming collagens (IV, VIII, X), which characteristically generate noncollagenous C-terminal domain dimers and N-terminal 7S domain tetramers; and membrane-anchored collagens (MACITs: XIII, XVII, XXIII, XXV) [19]. Among these types, COLI, COLIII, and COLV are mainly produced by fibroblasts, while COLIV is predominantly expressed by epithelial and endothelial cells. Notably, cancer cells and tumor-associated macrophages also produce collagen under some circumstances [20, 21].
Cancer cells influence collagen formation
During the occurrence and development of cancer, the ECM undergoes structural changes. In cancer cells, the content and distribution of collagen is modified to further coordinate cancer cell biological properties, including various gene mutations, transcription factors, signal transduction pathways, and receptors.
The heterogeneity of mutated genes is one of the major promoters for cancer cell behavior and influences the interaction between cancer cells and ECM components. The mutation of oncogenes, which are mainly divided into tumor suppressor genes and proto-oncogenes, also alters the collagen conditions in the tumor matrix.
The content and architecture of collagen are strongly altered by mutated tumor suppressor genes in cancer cells. The p53 pathway regulates the formation of tumor-associated collagen signature-3, which is referred to as a collagen bundle angled 60° to 90° relative to the cancer border and is indicated by the proliferation and invasion of cancer [22]. Mutated p53 in cancer cells, along with the activation of Janus kinase 2-signal transducers and activators of transcription (STAT) 3 signaling, influences the collagen production response to paracrine stimulation from pancreatic stellate cells [23]. In addition, the effects of collagen resolvents are associated with p53. The extracellular collagen-derived antiangiogenic factor Arresten, which is located in the C-terminal noncollagenous domain of COL4A1, has been linked to p53 activation [24]. The p53 gene upregulated collagen prolyl-hydroxylase to potentiate the production of full-length COL4A1, further enhancing the content of Arresten [25]. Collagen closely interacts with not only p53 but also other tumor suppressor genes associated with cancer processes. Cancer progression can be regulated by deleting a single copy of the phosphate and tension homology deleted on chromosome ten (PTEN) gene or by completely silencing this gene, resulting in the increased recruitment of cancer-associated fibroblasts (CAFs) and production of COL1A1 [26, 27].
Consistently, mutated proto-oncogenes combine with collagen to support cancer progression. Mutant Kras together with the epithelial-mesenchymal transition (EMT) regulator Snail enhanced collagen production by pancreatic cancer stellate cells, and silencing Kras expression markedly decreased COLI deposition in renal fibrosis [28, 29].
Transcription factors can lead to aberrant target gene expression and tumorigenesis, and nuclear factor kappa-B (NF-κB) and STATs mostly participate in collagen expression and organization. For example, COL2A1 was shown to be under the transcriptional control of the NF-κB subunit p65 in sarcomatous [30]. Collagen fibers showed less parallel alignment, less skewed distribution, and more direction variation rather than decreased numbers following combination treatment with the Janus kinase/STAT3 inhibitor AZD1480 and gemcitabine for pancreatic cancer [31].
Cancer cells further communicate with collagen via signaling pathways during the processes of cellular metabolism, proliferation, differentiation, and apoptosis. Transforming growth factor-β (TGF-β)/Smad signaling is a typical component of serine/threonine kinase signal transduction. Accumulating studies have revealed a positive role of TGF-β/Smad signaling in collagen modification. The architecture and mechanics of collagen fibers adjacent to epithelial lesions, rather than abundant bulk collagen, transformed the pancreatic epithelium into stiff fibrotic tissue via nonfunctional Smad4-phosphorylated myosin light chain 2 [32]. TGF-β sometimes reverses cancer cell functions via collagen. Collagen stiffness induced melanoma differentiation through the Yes-associated protein (YAP)/pax3/microphthalmia-associated transcription factor (MITF) axis, but in the presence of fibroblasts, TGF-β suppressed YAP/pax3/MITF expression and induced YAP/transcriptional enhanced associate domain/Smad-driven transcription, leading to dedifferentiation [33].
Other signaling pathways also affect collagen within cancer cells. Crosstalk between TGF-β and the Ras-Raf-mitogen-activated protein kinase (MEK)-extracellular signal-regulated kinase (ERK) signaling pathway increased collagen synthesis along with p38 activation in melanoma cells to promote cancer progression [34]. Overexpression of tRNAMeti increased the production of collagens and collagen-processing enzymes, especially COLII, forming a protumorigenic ECM [35].
Tyrosine kinase receptors are one category of principle collagen-related receptors that are expressed in various cancers. Fibroblast growth factor receptor (FGFR) 4-R388, in which Gly388 in the FGFR4 transmembrane domain was replaced with arginine, regulated the degradation of COLI, COLII, and COLIV by increasing MMP-14 protein expression in prostate cancer cells, especially within the tumor and in the fibrous capsule around the cancer [36]. The effect of epidermal growth factor receptor (EGFR) on collagen remains to be further studied. Collagen was reduced in recurrent breast cancer by combinatorial treatment with FGFR- and EGFR-specific inhibitors, similar to the effect of this treatment on primary tumors [37]. In contrast, c-Met expression, rather than EGFR expression, colocalized with abundant COLI in pulmonary adenocarcinoma [38].
The G protein family receptors, especially small G proteins, including Ras and Rho members, are important in collagen fiber properties and production. G proteins can promote matrix stiffness due to their collagen alignment change. The high stiffness increases nuclear localization of the transcription factor Twist1 by further reducing the expression of the cytoplasmic binding partner Ras-GTPase-activating SH3 domain-binding protein 2 to induce cancer EMT, invasion, and metastasis [39]. Cell division cycle 42, a member of the Rho family, was shown to regulate the thickness and contractility of collagen with the activation of MMP-9 [40]. Rho-associated coiled-coil kinase (ROCK)/Rho signaling may communicate with collagen directly through fibroblasts to regulate cancer cell behavior. At least two interconvertible types of cancer cell migratory motility were shown to be regulated by adhesion to collagen: mesenchymal motility was dependent on integrin and MMPs with Ras-related C3 botulinum toxin substrate 1 (RAC1) signaling and caused cells to appear elongated and bipolar, while amoeboid motility was dependent upon the ROCK/Rho kinase and caused cells to appear round, further leading to myosin-II light chain phosphorylation and actomyosin shrinkage [41]. The ROCK/Rho signaling pathway also influences collagen by other mechanisms. Acetylation of the COL1A1 gene promoter was facilitated by ROCK/Rho signaling pathways in breast cancer cells [42]. In addition, in pancreatic ductal adenocarcinoma (PDAC), collagen impairment via ROCK inhibition was independent of changes in fibroblast proliferation and survival [43]. Notably, the three‐dimensional collagen matrix was remodeled by PDAC cells, possibly by the fusion of ROCK with estrogen receptor (ER) causing increases in MMP-10 and MMP-13 [44].
The influence of collagen on cancer cell behavior
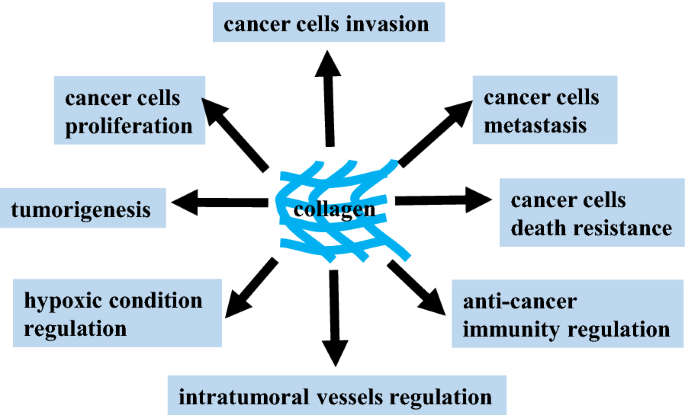
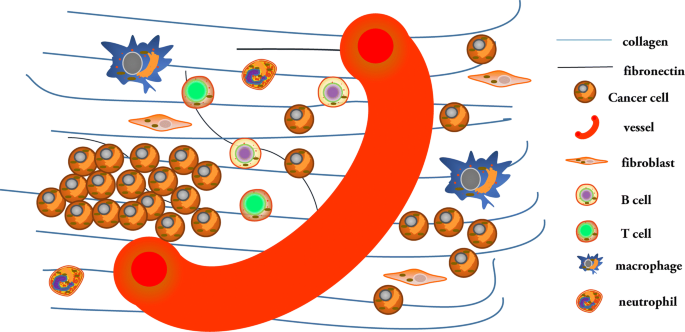
Cellular behavior is controlled by cell signal transduction pathways. Cells accept external signals through receptors and transmit them by cascade, which then transform extracellular signals into intracellular signals, causing physiological cellular reactions that regulate biological activities. Collagen, a component of the ECM, also influences cancer cell behavior (Fig. 1). Cancer cells reversely reshape collagen to form a reinforcing cell-collagen loop, which gradually fosters cancer progression.
Fig. 1
Collagen interacts with cancer cells mainly by directly connecting to cancer cell receptors. Discoidin domain receptors (DDRs) are a subfamily of tyrosine kinases that are divided into homologous DDR1 and DDR2 receptors. Collagens closely associate with preferred DDRs, such as COLIV with DDR1 and COLII and COLX with DDR2. A COLIV-DDR1-MMP-9-COLIV feed-forward loop was shown to promote the migration and adhesion of myeloid leukemia cells in bone marrow by activating AKT [45]. DDR1b phosphorylated at Tyr513 by COLI, as opposed to DDR1a, interacted with the signaling adaptor Src homolog 1 to affect focal adhesion kinase (FAK)-related protein-tyrosine kinase, resulting in N-cadherin upregulation in both primary and metastatic PDAC cells to induce EMT [46]. Others further reported that COLI activated DDR2 rather than integrin or TGF-βR to stimulate ERK2 in a Src-dependent manner; activated DDR2 then phosphorylated Snail1 at S82 and S104 and inhibited glycogen synthase kinase (GSK) 3β activity, ultimately contributing to sustained MMP-14 and collagen synthesis in breast cancer [47]. Nevertheless, DDR2 activated by collagen was not conducive to Src homolog 2 domain phosphorylation [48]. Notably, the binding of COL11A1 to both α1β1 integrin and DDR2 to activate the Src-phosphatidylinositol 3-kinase (PI3K)/AKT-NFκB signaling pathways induced the expression of three cisplatin-induced apoptosis inhibitors in ovarian cancer [49].
Adhesion between collagen and cancer cells, such as the adhesion of COLI and COLIV to cancer cells, impacts cancer progression [50]. The cadherin family represents one typical cell adhesion molecule. COLI stimulated E-cadherin upregulation to facilitate the migration of PDAC cells [51]. However, other studies on PDAC have reported the opposite effect of COLI on E-cadherin via different signaling pathways; Smad-interacting protein 1, a member of the small Zfh-1 family that acts as a transcriptional repressor, was induced by COLI to downregulate E-cadherin by simultaneously binding to two defined DNA target sites at E-boxes of the E-cadherin promoter through two zinc-fingers clusters [52]. The COLIV-regulating chemokine (C-C motif) ligand (CCL) 5 and CCL7 were associated with the alteration of E-cadherin to influence EMT, further promoting liver metastasis [53]. COLIV not only promoted a decrease in E-cadherin expression, an increase in N-cadherin expression, and upregulation of Snail1, Snail2, and Sip1 (E-cadherin transcriptional repressors that bind at E-boxes of the E-cadherin promoter) but also induced FAK and ERK1/2 activation in affiliation with TGF-β during EMT, resulting in increased MMP-2 secretion and enhanced cell migration [54]. In addition, the mediation of prostate cancer metastasis by COLXXIII corresponds to changes in OB-cadherin, α-catenin, β-catenin, γ-catenin, vimentin, and galectin-3 protein expression [55].
Integrin, a typical adhesion molecule in cancer cells, often mediates cancer cell behavior, especially when combined with collagen. Integrin comprises two units: α and β. Different types of collagen bind to various integrins in numerous signaling pathways in cancer cells. The binding of integrin to collagen led to the activation of AKT/PI3K signaling, mitogen-activated protein kinase (MAPK) signaling, and Rho family signaling, and the MEK/ERK signaling pathway especially regulated αv integrin subfamily members such as αvβ3 and αvβ5, inducing the proliferation and invasion of squamous cell carcinoma (SCC) cells [56]. Additionally, the deposition of collagen through integrin-regulated ROCK, FAK, and AKT activation inactivated GSK3β and increased the nuclear localization of the mechanotranscription coactivator β-catenin to promote cutaneous SCC progression [57]. Various experiments have further revealed the effects of specific types of collagen in combination with integrin on cancer. COLI is a typical interstitial matrix collagen via integrin to induce cancer cell behavior. Remodeled COLI affected the invasion of ovarian cancer cells by mediating the integrin-PTEN/PI3K/AKT signaling pathway [58]. COLI and α2β1 integrin-promoting cathepsin B-mediated invasiveness were associated with secreted acidic and cysteine-rich proteins in melanoma [59]. The mediation of COLI by α2 integrin led to EMT-like changes, such as downregulated E-cadherin and β-catenin expression, decreased differentiation, increased clonogenicity, and increased colorectal cancer stem cells [60]. The expression of αv integrin response to COLI was enhanced by melanoma cells to promote the upregulation of protein kinase C (PKC) α, thereby relocating endogenous p53 protein [61]. During its adhesion to COLI, mda-9/syntenin at the plasma membrane facilitated processes in the formation of β1 integrin signaling complexes, including the assembly of the integrin-linked kinase (ILK)-PINCH1-α-parvin complex and its translocation to the cell membrane, leading to the activation of AKT, RAC1, and ERK1/2 to promote cancer metastasis [62]. The linearization and matrix compaction of COLI, via β1 integrin-FAK signaling modulated myosin IIA, was exhibited by most radiation-induced breast cancer cells [63]. COLIV accounts for the basement membrane. The high-density COLIV matrix induced the formation of cancer cell invadopodia and actin-rich proteolytic protrusions, which locally degraded collagen via αvβ3 integrin [64, 65]. The COLIV/β1 integrin signaling pathway significantly stimulated Src and ERK phosphorylation, reducing cell stiffness and accelerating cell motility [66]. Ras GTPases, Rac GTPases, PI3K, and PKC participated in melanoma cell migration mediated by COLIV/β1 integrin [67]. In soft-tissue sarcoma, the interplay of COLVI and NG2 triggered PI3K activation through α2β1 integrin, which was associated with adhesion, survival, aggregation, and migration and did not directly influence cell mitosis [68]. Both matrix collagen and basement membrane collagen communicate with integrin to impact cancer cell behavior, and numerous other collagens also bind to integrin to regulate cancer progression. COLXIII in breast cancer [69], COLXVI in glioblastoma [70], and COLXVI in OSCC [71] induce β1 integrin to promote cancer stemness, invasion, and drug resistance. Even collagen glycosylation modulates integrin binding. Galactosylation occurred on the periphery of α2β1 integrin, where it interacted with α1(IV)382–393 but occurred in the middle of α3β1 integrin, where it interacted with α1(IV)531–543 in melanoma cell adhesion [72].
Collagen can stimulate additional signaling pathways in cancer cells to exert various functions. The increased expression of COL1A1 affected the caspase-3/PI3K/AKT pathways to inhibit cell apoptosis in cervical cancer tissues [73]. After the withdrawal of rapamycin treatment, mutated COL1A1 reinforced PI3K–AKT-mammalian target of rapamycin (mTOR) signals in cancer stem cells to sustain the metastatic burden of ERα-positive breast cancer cells; however, lung metastases were independent of mTOR signaling [74]. In addition, increased COLI did not alter primary tumor growth and ERα expression but enhanced circulating cancer cells and metastasizing cancer cells with decreased phospho-STAT5 expression, increased phospho-ERK1/2, and increased phospho-AKT expression; this phenomenon coincided with the formation of invasive protrusions of the primary tumor harboring collagen fibers angled perpendicularly to the tumor mass [75]. However, COLI in non-small cell lung cancer (NSCLC) induced mTOR activation through an AKT-independent pathway, leading to EGFR-tyrosine kinase inhibitor resistance [76]. The Notch3-COL4A2 loop promoted anoikis resistance with a reduction in phosphorylated AKT and ERK 1/2 in ovarian cancer cells [77]. Although both collagen glycation and carbamylation affected the metastasis of cancer cells, glycation caused a more obvious delay in cell adhesion time and deficient actin stress fibers and inhibited the mean cell speed and FAK phosphorylation state more than carbamylation [78]. However, increased collagen in fibrosarcoma tissue inhibited tumor growth and metastasis because the tumor necrosis factor (TNF) receptor 2/p38 MAPK signaling pathway activated collagen expression via gadolinium-containing fullerenol [79]. This distinction implies that different cancer cells facilitate collagen expression to exert inverse effects on cancer progression.
The relationship among exosomes, microRNAs and collagen in cancer
Recent studies have highlighted the relationship among exosomes, microRNAs (miRNAs) and collagen in cancer [80,81,82].
MiRNAs are a class of small, noncoding RNAs that act as epigenetic regulators of various physiological and pathological processes [83, 84]. The deregulation of miRNAs is associated with the initiation and progression of many diseases, such as cardiovascular diseases, infectious diseases, diabetes, central nervous system-related diseases, and cancer [85,86,87,88]. MiRNAs exert their regulatory activity by affecting a variety of physiological and pathological processes in cancer. Several studies have documented the roles of miRNAs in tumor growth, angiogenesis, and metastasis [89, 90], as well as their utility as diagnostic and therapeutic biomarkers [91,92,93]. Regarding the influence of miRNAs on collagen, various miRNAs are regulated in cancer cells to affect distinct functions. Breast cancer cells downregulate miR-196b-5p, which decreases COL1A1 levels, to induce growth and metastasis [94]. Intestinal gastric cancer cells suppress COL1A2 via miR-25 to promote EMT and angiogenesis [95]. MiR-27b-3p and miR-455-3p enhance cancer cell quiescence in response to activated p53 to increase drug resistance and recurrence [96]. The miRNA let-7d inhibits pancreatic stellate cell activation [97] and macrophage infiltration by targeting COL3A1 in renal cell carcinoma [98]. Collagen-related enzymes are also differentially regulated by miRNAs in cancer cells. MiRNA-29a targets heat shock protein 47 in cervical SCC [99], and miR-26a/b affects lysyl oxidase-like (LOXL) 2 and procollagen-lysine, 2-oxoglutarate 5-dioxygenase 2 (PLOD2) levels in renal cancer cells [99]. Losartan improves the efficacy of chemotherapy in ovarian cancer partly by inducing antifibrotic miRNAs to normalize the ECM [100].
Exosomes are membrane-enclosed structures that facilitate communication between cancer cells and the ECM to influence cancer cell survival, growth, and metastasis and the immune system [101]. Cancer-derived exosomes induce the formation of CAFs in the collagen matrix to promote EMT [102] and increase the secretion of MMP-14 to regulate collagen [103]. In addition, collagen enhances exosome secretion [104]. Collagen and exosomes form a mutually beneficial feedback loop to promote cancer progression.
The reciprocity between collagen and cancer cells under hypoxic conditions
Hypoxic conditions, a common condition in collagen-rich ECM, intensify cancer progression based on the interaction between cancer cells and collagen. Hypoxia-inducible factor (HIF)-1, LOX, and MMP participate in the process. COLI fibers are reduced in the hypoxic microenvironment via increased expression of LOX in renal cell carcinoma, decreased expression of MMP-3 in breast cancer, or decreased expression of MMP-1 and MMP-16 in prostate cancer [105, 106]. Collagen can be remodeled by HIF-1 regulating LOXL1, LOXL2, and LOXL4 [107].
Other substances, such as hepatitis transactivator protein X in liver cancer [108], LKB1 via COLIV and β1 integrin in lung cancer [109], and HIF1α-dependent PLOD2 in primary sarcomas and pulmonary metastasis [110], participate with collagen in the HIF/LOX pathway. Hypoxia increased the expression of procollagen-lysine as collagen crosslinker to further enhance collagen fiber size and thus promote cancer metastasis [111]. COLI fibers exhibited covalent crosslinking with prolyl 4-hydroxylase alpha 1 and prolyl 4-hydroxylase alpha 2 in differentiated cell types of triple-negative breast cancer, and the prolyl 4-hydroxylase alpha 1/HIF-1 axis increased chemoresistance [112, 113]. Even mutant p53 regulated the expression of COL7A1 in NSCLC, not by influencing HIF-1 binding to DNA, but rather by inhibiting its transcriptional activity [114].
Hypoxic conditions are closely associated with vessels in cancer. Matrix collagen stiffness, rather than collagen density, alters vascular growth and integrity. Moreover, increased fibril density decreased vessel network formation, while increased interfibril branching improved vessel volume density and formation, which were markedly dependent on the changed temporal and spatial depositions of COLIV [115]. In addition, neovessel branching is associated primarily with collagen crosslinking rather than with collagen content [116]. Notably, the von Hippel–Lindau protein directly regulated COL4A2 assembly independent of HIF-α to restrain cancer angiogenic formation [117]. Further reflecting the relationship between collagen and cancer cells with regard to treatment, improved tumor stromal pO2 levels and the recovery of blood flow were associated with the content and diameter of collagen fibril under imatinib treatment [118].
Interaction between collagen and tumor matrix components
The complex interactions among collagen and matrix proteins within ECM cells contribute to cancer initiation and progression, as shown in Fig. 2.
Fig. 2
Collagen in cancer tissue accompanies various categories and degrees of myeloid-lineage immune cells, including mast cells, macrophages, and neutrophils, and lymphocytes, including T cells and B cells, to reflect different cancer progression stages [119]. For example, pancreatic cancer stromal compositions were classified as inert, dormant, fibrogenic, or fibrotic based on α-smooth muscle actin (SMA) and COLI expression; these differing stromal compositions individually influenced the levels of CD4+ T cells, CD8+ T cells, macrophages, and neutrophils [120]. Increased infiltration of macrophages and lymphocytes was also observed in subcutaneous adipose collagen in gastrointestinal cancer patients with cachexia [121]. However, increased stromal COL10A1 was accompanied by low numbers of total tumor-infiltrating lymphocytes in ER-positive/EGFR2-positive breast cancer [122].
The location and quantity of collagen with lymphocytes reinforce discrepant cancer progression via distinct signaling pathways. A high-density collagen matrix, rather than a low-density matrix, decreases T cell abundance and cytotoxic T cells but upregulates regulatory T cells [123, 124]. Aligned fibers and collagen density around vascular regions and epithelial cancer cells restricted the migration of T cells and limited their entry into the tumor mass [125]. Moreover, others have further reported that the distribution of collagen with lymphocytes affects cancer progression, but the effects on different cancers remain distinct. COLI was not associated with T cell deficiency in PDAC [126]. However, others noted that the metastatic urothelial cancer response to programmed death-1 and programmed death-ligand 1 treatment involved the movement of CD8+ T cells from the cancer parenchyma to the collagen-rich peritumoral stroma [127]. Nevertheless, contact between infiltrating T cells, Tregs and antitumor T cells with collagen was inhibited by members of the TGF-β pathway to provide antimelanoma-immunity [128, 129].
Furthermore, both adaptive immunity and innate immunity affect collagen. Collagen deposition and linearization are positively related to macrophage activity [130]. Additionally, denatured collagen acts as a strong chemoattractant for macrophages that mediate the promotion of cancer [131]. The possible reasons for these functions of collagen are described below. Macrophages closely interact with stellate cells to influence the collagen matrix [132]. Tumor-associated macrophages orchestrate the deposition, crosslinking, and linearization of collagen fibers, specifically COLI, VI, and XIV, at areas of tumor invasiveness [133]. Although macrophages do not exhibit efficient collagen internalization of mesenchymal origin, those originating from circulating CCR2 monocytes internalize collagen in an MMP-dependent manner, which is mediated by the mannose receptor rather than by β1 integrin [134,135,136]. In the collagen-rich breast cancer environment, coexpression of fibroblast activation protein and heme oxygenase-1 in macrophages resulted from an early innate regenerative response to IL-6 to directly facilitate transendothelial migration [137]. Cyclooxygenase-2 also induced overall collagen deposition and macrophages in early-stage breast cancer [138]. Notably, in the context of an αvβ3 integrin-specific collagen hydrogel, M2-like immunosuppressive macrophages promoted angiogenesis in glioblastoma, while M1-like proinflammatory macrophages suppressed angiogenesis, which was regulated by Src-PI3K-YAP signaling associated with TGF-β1 [139]. IL-10 induces M2 differentiation in heterospheroid macrophages [140]. In contrast, collagen degradation is enhanced not by M2 macrophages but rather by M1 macrophages, as endogenous macrophages are recruited, modified and activated to produce MMPs and hepatic growth factors, thereby enhancing hepatocyte proliferation and the release of TNF-related apoptosis-inducing ligands by natural killer cells, leading to hepatic stellate cell apoptosis [141]. Moreover, there are other important innate immune cells in the ECM that exhibit anticancer immunity or immunosuppressive effects. Collagen deposition can be induced by neurofibromatosis type 1-recruited mast cells, which mediate stem cell factor/c-kit signaling in neurofibromas [142]. Collagen-dense tumors increased granulocyte monocyte-colony stimulating factor, which was associated with neutrophil signaling [143]. However, COLXVII overexpression in cancer cells by activated leukocyte-associated immunoglobulin-like receptor-1 diminished natural killer cell cytotoxic activity [144]. By interaction with collagen in ovarian and breast cancer, tumor-associated dendritic cells expressed the highly receptor CD305/leukocyte-associated immunoglobulin-like receptor-1 [145].
Fibroblasts are another important cell type in the ECM. CAFs cause various collagen deposits to develop into intratumoral fibrosis, resulting in cancer occurrence, differentiation, and invasion [4]. These CAFs can even originate from adipose tissue-derived stromal cells [146]. CAFs are classified as tumor-restraining, tumor-promoting, secretory, or ECM-remodeling cells [147], which also exhibit CAF functions via mutated genes [148], the secreted cytokines IL-1β, TNF-α and NF-κB, inflammatory signals, epigenetic regulation, etc. [6]. Cancer cells can invade collagen with fibroblasts, which are characterized by cell cycle phases and small cancer cell nest formation [149]. Additionally, CAFs exert anticancer and cancer-induced effects to show dual influences for cancer progression [150].
Some ECM proteins, mainly including MMPs, hyaluronic acid, fibronectin, and laminin, regulate cancer cell invasion and migration with collagen. The functions of MMPs with collagen are summarized in Table 1, and MMP activity with contractility necessarily establish the ECM stiffness associated with collagen bulk and distribution [151]. The collagen gel storage modulus with glycosaminoglycans chondroitin sulfate and hyaluronic acid was dependent on both the fiber diameter and network mesh size [152]. When cancer cells encountered stiff collagen fibers at fibronectin-rich invasive fronts, they engaged αvβ1 integrin to recruit vinculin and zyxin to focal adhesions sites in a tension-dependent manner to induce PI3K signaling [153]. Similarly, with increased COLI in the laminin-rich ECM regarded as an early tumor microenvironment, cancer stem cells maintained their endothelial-like gene signatures and secreted high levels of VEGFR-2 in a paracrine and autocrine manner to simulate progression [154]. In contrast, unlike the laminin-rich microenvironment, a COLI Matrigel was sufficient to induce colon cancer mesenchymal gene expression, suppressing hepatocyte nuclear factor 4α and its target genes [155]. Additionally, CAFs initially constructed the ECM network by depositing fibronectin, followed by the preferential interaction of COLI with relaxed fibronectin, which, in turn, limited the stretching and mechanical unfolding of fibronectin, leading to collagen superseding fibronectin [156, 157]. Other types of collagen also have functions. The expression of COLIV was associated with the expression of fibronectin and laminin during central nervous system metastasis [158]. The binding of COLXV, rather than I, III, IV, and V, to fibronectin, laminin, and vitronectin inhibited the adhesion and migration of fibrosarcoma cells [159].Table 1 Pathological functions of MMPs associated with collagen in cancer
Various experiments and clinical data have revealed collagen to be a prognostic factor correlated with cancer differentiation, cancer invasion, lymph node metastasis, and clinical stage in cancer patients (Table 2). Lower COLI and COLIV content is a biomarker of differentiated tumors and proliferation potency of cancer cells [160, 161]. The increased expression of COLIV and COLVI and the collagen structure reflect tumor angiogenesis and glioblastoma progression [162]. Even hypomethylation of the COL17A1 promoter is associated with advanced stage, increased invasion of breast cancer, lung adenocarcinoma, cervical cancer, neck SCC, and lung SCC [163].Table 2 Collagen as a prognostic factor for cancer patients
Factors other than collagen content are correlated with clinical outcome [164], such as collagen alignment and distribution, which also affect cancer. During tumor progression, collagen exhibits different signatures. Tumor-associated collagen signature 1 (TACS-1) indicates the presence of dense collagen near the cancer, TACS-2 represents collagen fibers parallel to the tumor edge, and TACS-3 depicts radially aligned collagen fibers [165]. TACS-3 was related to cancer cell invasion and poor survival in breast carcinoma and in situ breast ductal carcinoma [166, 167]. TACS-3 was shown to be driven by increased plasminogen activator inhibitor 1 via ERK signaling and promoted the migration of triple-negative breast cancer cells [168]. The elevated density and depth of collagen deposition showed high proliferation and invasion of cancer cells [169, 170]. Although COLI and COLIV are expressed in different tumor stroma compartments in pancreatic cancer tissue, they stimulate proliferation, migration, and antiapoptosis. The main form of COLI was generated by pancreatic stellate cells attributed to cancer cells in an indirect contactable desmoplastic area to activate TGF-β, while COLIV was produced by cancer cells themselves to form an autocrine loop in direct proximity to cancer cells, causing discontinuous basement membrane-like structures that interacted with the COLIV CB3 region and β1 integrin of the cancer cells [171]. However, another study showed that COLV was expressed by pancreatic stellate cells via paracrine loops in PDAC [172]. The desmoplastic reactions in primary cancer were divided into mature, intermediate, and immature based on the presence of keloid-like collagen and myxoid stroma; immature desmoplastic reactions were associated with higher T and N stages, more extensive liver metastasis, and higher recurrence rate than other reaction types [173]. Furthermore, COLVII positively regulates the abundance of the cell polarity markers E-cadherin and B-catenin [174]. The mechanisms underlying these important collagen properties are described below. Among five collagen parameters (alignment, density, width, length, and straightness), increased collagen width is the most powerful parameter for predicting cancer prognosis [175]. The elasticity of the collagen matrix is controlled by fibril bending stiffness rather than by fibril diameter or intrafibrillar crosslinking [176]. Increased collagen fiber alignment, elevated levels of immunoreactive glycosaminoglycans such as heparan sulfate and chondroitin sulfate, and decreased levels of the proteoglycan decorin enhance the stiffness of carcinoma tissues [177]. In contrast, others have notably reported that the distribution of collagen had no effect in the regenerated cervical tissue following excisional cervical intraepithelial neoplasia [178].
Collagen plays an important role in therapy resistance. In esophageal SCC, increased collagen content was associated with chemotherapy resistance via the MAPK and PI3K/AKT pathways [179]. Even at metastatic sites, collagen crosslinking increases tissue stiffness to promote resistance to treatment [180]. Furthermore, increased collagen content was accompanied by increased hyaluronan accumulation, contributing to doxorubicin drug resistance in pancreatic cancer [181]. Specifically, different collagen types exhibit distinct treatment resistances. COLI induced resistance to drugs, such as cisplatin and mitoxantrone, by activating β1 integrin followed by the FAK/PI3K/AKT pathway in ER-positive cancer cells, the MAPK pathway in triple-negative cancer cells, the coexpression of LOX with COL1A2 in ovarian cancer, and the TGF-β1/Smad3-mediated expression of COLI and COLIII in bromocriptine-resistant prolactinoma cells [182,183,184]. Other mechanisms of drug resistance include COLI-induced tau upregulation, resulting in paclitaxel resistance in ovarian carcinoma [185]. The resistance to radiation of renal cancer cells with intact COLI rather than micronized COLI was mediated by apoptosis attenuation rather than cell cycle redistribution via the PI3K/AKT pathway [186]. In addition, other collagens also play important roles in therapy resistance. COL3A1 optimally predicted the absence of a response to neoadjuvant treatment in rectal cancer and the resistance of ovarian cancer cells to topotecan and paclitaxel [187, 188]. High COLVI expression promoted anoikis resistance and affected the response of salivary gland cancer to radiotherapy and colorectal cancer to adjuvant chemotherapy [189,190,191]. The reactions of cancers to antiangiogenic therapy have also been closely associated with COLIV. The reduced binding of collagen to PDAC cell surface receptors promoted resistance to VEGF therapy via TGF-β signaling [192], but antiangiogenic therapy increased intratumoral adenovirus distribution by decreasing COLIV [193]. Another study showed that the correlation of bevacizumab with elevated tumor stiffness was driven by hypoxia, leading to increased hyaluronic acid and sulfated glycosaminoglycan contents without significantly changing collagen deposition [194]. COL11A1 induced chemoresistance and exerted antiapoptosis effects in ovarian cancer cells by mediating the transcriptional activation of NF-κB to upregulate the Twist family [195].
Collagen can also be used for the imaging of targeted sites. Imaging of the collagen content and arrangement is conducive to the assessment of tissue stiffness, metabolism, and drug resistance [196,197,198], and technology to utilize these data is currently under development [199]; for instance, magnetic resonance apparent diffusion coefficient values were used to indicate a negative correlation of collagen with esophageal SCC cells [200]. The label-free Raman spectroscopic measurements used to distinguish radiation-sensitive tumors were based on differences in collagen content [201].
Because collagen is closely associated with clinical outcome, it can also be used in clinical applications (Fig. 3). Collagen can be utilized as a predictor of prognosis and recurrence, in diagnosis, as a therapy resistance biomarker, as a targeted-therapy strategy, and as a drug carrier.
Fig. 3
Collagen-related therapy
Cancer resistance has substantially hindered the ability to control cancer. Therefore, both cancer cells and tumor microenvironment must be treated, and collagen is a potential target. Moreover, collagen has obvious genetic stability, and its spatial structure remains relatively stable. As collagen is basically present in various types of cancer, the treatment value of modifying collagen conditions in cancer is worth exploring.
Collagen can be regulated by different types of inhibitors against biosynthesized processes and distribution arrangements by interfering with collagen biosynthesis enzymes, disturbing cancer cell signaling pathways, mediating ECM components, or directly utilizing collagenases (Table 3). Although many types of inhibitors exist, most have been investigated in only cell and animal experiments. These indirect methods of modifying collagen are also partially attributed to other intricate related molecular mechanisms and signaling pathways. The treatment of collagen in the current study is mainly to target the function of CAFs. While quiescent CAFs are characterized by the presence of lipid droplets loaded with vitamin A in the cytoplasm, these perinuclear lipid droplets disappear and express the activation marker α-SMA after CAF activation [202]. Moreover, low expression of retinoid receptors is associated with poor survival of pancreatic cancer [203]. Therefore, the vitamin A analog all-trans retinoic acid in the treatment of PDAC induces CAF to be at quiescent status and inhibits cancer cell migration and EMT; however, the relevant clinical trials are only stage I in the recruitment state (NCT03307148) [204]. In addition, some studies focus on the interaction of cancer cells with collagen, which regulates collagen by targeting cancer cells. For instance, the development of PDAC is also associated with vitamin D, which affects insulin synthesis and secretion via reduced CYP24A1 in islets but impairs the anti-proliferation in transformed duct cells via increased CYP24A1 [205]. The vitamin D signaling pathway has the functions of anti-proliferation, pro-differentiation, anti-inflammation, pro-apoptosis, and immune regulation. There is also a stage III clinical trial recruiting to study the influence of high-dose vitamin D3 intake on pancreatic cancer surgery outcomes (NCT03472833). Moreover, vitamin D receptor ligand calcipotriol regulates CAFs to reprise the quiescent state [206], and paricalcitol was selected to participate in ongoing clinical trials on PDAC therapy (NCT03883919, NCT03415854). Hydroxychloroquine can also effectively inhibit the proliferation and metabolic activity of fibroblasts for cancerous interstitial fibrosis and inhibit cancer cells autophagy. However, the combination of gemcitabine hydrochloride and nab-paclitaxel plus hydroxychloroquine did not increase overall survival for metastatic pancreatic cancer; thus, the clinical effect still needs to be further explored [207]. Notably, although MMPs degrade collagen, they also induce cancer angiogenesis; thus, they have complex and paradoxical effects on cancer progression. The combination of an anti-MMP-9 antibody and nab-paclitaxel-based standard cytotoxic therapy was shown to decrease COLI and the metastatic burden compared with that achieved with nab-paclitaxel-based standard cytotoxic therapy in PDAC mouse models [208]. Moreover, collagen-targeted treatments have paradoxical effects on drug delivery and treatment efficacy within the same cancer type. In particular, collagenases monotherapy for cancer may have obvious side effects, and it may even exert opposite effects than those intended. Addition of the sonic hedgehog antagonist vismodegib to gemcitabine did not improve the survival outcomes of metastatic pancreatic cancer patients in a phase Ib/II trial [209]; in contrast, halofuginone disrupted collagen barriers to effectively deliver the drug and promote anticancer immunity [210]. While the antifibrotic drug pirfenidone was effective in early-stage liver fibrosis, it did not influence advanced liver fibrosis and initiation-promotion liver cancer [211].Table 3 Typical inhibitors and drugs that regulate collagen biosynthesized processes and collagen distribution arrangement in cancer studies
Combining collagen inhibitors and standard therapeutics, such as chemotherapy and radiotherapy, is a promising anticancer strategy. Collagenase combined with trastuzumab via thermosensitive hydrogels exerted an anticancer effect on animals [212]. Nitric oxide activated endogenous MMP-1 and MMP-2 to deplete collagen, and it significantly improved anticancer efficacy while exerting no overt toxicity in animal models [213]. Under hypoxic cancer conditions, hyperbaric oxygen therapy decreased collagen deposition to enhance chemotherapy efficacy and photodynamic upconversion nanophotosensitizer cancer therapy [214].
Preclinical studies on collagen-related therapy partly demonstrated encouraging outcomes. However, few clinical trials have been conducted, and most have focused on signaling pathways or receptors. These indirect collagen-related treatments remain controversial due to the potential effects of other mechanisms. For example, cilengitide inhibiting αvβ3 and αvβ5 integrins to influence the connection with cancer cells and collagen did not show evident clinical benefits [215]. There are major drugs of direct collagen depletion or collagen alignment changes, but their effects are not clear and need to be further explored (Table 4). Confusing and conflicting results for anti-collagen therapy suggest complex collagen properties. As revealed herein, physiological, pathological and clinical analyses reveal the dual roles of collagen. Collagen has essential functions in normal tissues but also plays important roles in cancer progression and clinical outcomes. Collagen depletion results in the activation of residual cancer cells and incomplete ECM and microvessels. Collagen can function as a barrier for certain stages of cancer, but it can also enhance other stages of cancer. In addition, although collagen can be degraded, its decomposition products can continue to function and thus promote cancer angiogenesis and invasion. The interaction of collagen with other components in the ECM also shows dual effects on cancers. CAFs, which mainly produce stromal collagen, express anti-tumor and tumor-promoting effects; epithelial and endothelial cells, which mainly produce the collagen of basement membrane, can not only maintain vascular stability but also promote cancer angiogenesis and contribute to cancer cells penetration into blood vessels. The individual differences, genetic heterogeneity, and epigenetic heterogeneity of cancer patients can also affect the efficacy of drugs in clinical trials. Therefore, balancing the content, crosslinking, alignment, and distribution of collagen may be a reasonable strategy for cancer treatment. Nanoparticles, nanoplatforms, and nanoenzymes exhibit the expected gratifying properties.Table 4 Collagen-targeted agents directly influencing collagen content and distribution in clinical trials for cancers
Nanoparticles influence the use of collagen for anticancer therapy in animal models, including combinations with other chemotherapy drugs and treatment regimens [216]. Losartan nanoparticles significantly decrease collagen content to improve tumor penetration and tumor treatment efficiency [217], and losartan in combination with photodynamic nanoplatforms suppressed tumor volume in a breast cancer mouse model [218]. The implantation of nanoparticle/losartan-loaded hydrogel enhanced the intratumoral distribution and anticancer effect of nanoparticles in mice [219]. In addition, supplementation with other nanoparticles further orchestrated collagen normalization rather than destruction, thereby improving the survival rates of cancer patients [220]. A near-infrared light irradiation-activated semiconducting polymer nanoenzyme efficiently digested collagen, leading to nanoparticle accumulation in cancer tissue and the consequential improvement of photothermal therapy [221, 222].
Collagen can also act as a drug carrier or a drug targeted site. Hybrid collagen-cell penetrating peptide carriers improved resistance to enzymatic degradation [223]. Treating cancer with the single-chain fragment of cetuximab along with its collagen-binding domain demonstrated effective results [224]. Collagen affinity can also be used to mediate targeted immunotherapy antibodies. Fusion of the collagen-binding protein lumican and cytokines increased the efficacy of systemic immunotherapies in a melanoma model [223]. The therapeutic use of immune checkpoint inhibitors and interleukin-2 by conjugation (for antibodies) or recombinant fusion (for cytokine) to the von Willebrand factor A3 domain (a collagen-binding domain) eradicated tumors and exhibited obvious safety and efficacy in a breast cancer model [225]. Cancer-collagen-targeting immunoconjugate therapy was evidently applicable to anticancer therapy [226].
Conclusions
Cells and molecules in the tumor microenvironment have dual effects on cancer progression. The role of collagen is a double-edged sword in cancer. On the one hand, collagen, cancer cells, other cells, and other matrix molecules mutually form an inter-reinforcing loop. This loop contributes to the development of cancer by inducing cancer cells proliferation, migration, and metastasis. On the other hand, preclinical and clinical studies have demonstrated that collagen may slow the development of cancer cells to some extent under some conditions. In summary, the association of collagen with cancer is only partially understood, and future studies are needed to elucidate detailed collagen biological mechanisms in cancer tissue that can be applied to precisely regulate collagen balance to achieve the maximum benefit of treatment. This new strategy combined with other treatment modalities can ultimately improve patient survival and quality of life.
Availability of data and materials
Not applicable
Abbreviations
CAFs:
cancer-associated fibroblastsCCL:
chemokine (C-C motif) ligandCOL:
collagen typeDDR:
discoidin domain receptorECM:
extracellular matrixEGFR:
epidermal growth factor receptorEMT:
epithelial–mesenchymal transitionER:
estrogen receptorERK:
extracellular signal-regulated kinaseFACITs:
fibril-associated collagens with interrupted triple helicesFAK:
phosphate and tension homology deleted on chromosome tenRAC1:
Ras-related C3 botulinum toxin substrate 1SCC:
squamous cell carcinomaSMA:
smooth muscle actinSTAT:
signal transducers and activators of transcriptionTACS:
tumor-associated collagen signatureTGF-β:
transforming growth factor-βTNF:
tumor necrosis factorYAP:
Yes-associated protein
References
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424.ArticlePubMedGoogle Scholar
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69:7–34.ArticlePubMedGoogle Scholar
Lambrechts D, Wauters E, Boeckx B, Aibar S, Nittner D, Burton O, Bassez A, Decaluwe H, Pircher A, Van den Eynde K, et al. Phenotype molding of stromal cells in the lung tumor microenvironment. Nat Med. 2018;24:1277–89.ArticleCASPubMedGoogle Scholar
Discher DE, Smith L, Cho S, Colasurdo M, Garcia AJ, Safran S. Matrix mechanosensing: from scaling concepts in ‘Omics data to mechanisms in the nucleus, regeneration, and cancer. Annu Rev Biophys. 2017;46:295–315.ArticleCASPubMedPubMed CentralGoogle Scholar
Whatcott CJ, Diep CH, Jiang P, Watanabe A, LoBello J, Sima C, Hostetter G, Shepard HM, Von Hoff DD, Han H. Desmoplasia in primary tumors and metastatic lesions of pancreatic cancer. Clin Cancer Res. 2015;21:3561–8.ArticleCASPubMedPubMed CentralGoogle Scholar
Gu L, Shan T, Ma YX, Tay FR, Niu L. Novel biomedical applications of crosslinked collagen. Trends Biotechnol. 2019;37:464–91.ArticleCASPubMedGoogle Scholar
Ishikawa Y, Rubin K, Bachinger HP, Kalamajski S. The endoplasmic reticulum-resident collagen chaperone Hsp47 interacts with and promotes the secretion of decorin, fibromodulin, and lumican. J Biol Chem. 2018;293:13707–16.ArticleCASPubMedPubMed CentralGoogle Scholar
Shi R, Hu W, Zhang Y, Gao S, Smith AH, Ye J, Cai L, Graham LM, Li C. Ascorbate inducible N259 glycans on prolyl 4-hydroxylase subunit alpha1 promote hydroxylation and secretion of type I collagen. Cell Mol Life Sci. 2019;76:3449–64.ArticleCASPubMedPubMed CentralGoogle Scholar
Elia I, Rossi M, Stegen S, Broekaert D, Doglioni G, van Gorsel M, Boon R, Escalona-Noguero C, Torrekens S, Verfaillie C, et al. Breast cancer cells rely on environmental pyruvate to shape the metastatic niche. Nature. 2019;568:117–21.ArticleCASPubMedPubMed CentralGoogle Scholar
Scietti L, Chiapparino A, De Giorgi F, Fumagalli M, Khoriauli L, Nergadze S, Basu S, Olieric V, Cucca L, Banushi B, et al. Molecular architecture of the multifunctional collagen lysyl hydroxylase and glycosyltransferase LH3. Nat Commun. 2018;9:3163.ArticlePubMedPubMed CentralCASGoogle Scholar
Bekhouche M, Colige A. The procollagen N-proteinases ADAMTS2, 3 and 14 in pathophysiology. Matrix Biol. 2015;44–46:46–53.ArticlePubMedCASGoogle Scholar
Tjin G, White ES, Faiz A, Sicard D, Tschumperlin DJ, Mahar A, Kable EPW, Burgess JK. Lysyl oxidases regulate fibrillar collagen remodelling in idiopathic pulmonary fibrosis. Dis Model Mech. 2017;10:1301–12.ArticlePubMedPubMed CentralGoogle Scholar
Gautieri A, Passini FS, Silvan U, Guizar-Sicairos M, Carimati G, Volpi P, Moretti M, Schoenhuber H, Redaelli A, Berli M, Snedeker JG. Advanced glycation end-products: mechanics of aged collagen from molecule to tissue. Matrix Biol. 2017;59:95–108.ArticleCASPubMedGoogle Scholar
Scarpellini A, Huang L, Burhan I, Schroeder N, Funck M, Johnson TS, Verderio EA. Syndecan-4 knockout leads to reduced extracellular transglutaminase-2 and protects against tubulointerstitial fibrosis. J Am Soc Nephrol. 2014;25:1013–27.ArticleCASPubMedGoogle Scholar
Van Doren SR. Matrix metalloproteinase interactions with collagen and elastin. Matrix Biol. 2015;44–46:224–31.ArticlePubMedCASGoogle Scholar
Ohlund D, Lundin C, Ardnor B, Oman M, Naredi P, Sund M. Type IV collagen is a tumour stroma-derived biomarker for pancreas cancer. Br J Cancer. 2009;101:91–7.ArticleCASPubMedPubMed CentralGoogle Scholar
Qiu S, Deng L, Liao X, Nie L, Qi F, Jin K, Tu X, Zheng X, Li J, Liu L, et al. Tumor-associated macrophages promote bladder tumor growth through PI3K/AKT signal induced by collagen. Cancer Sci. 2019;110:2110–8.ArticleCASPubMedPubMed CentralGoogle Scholar
Kenny TC, Schmidt H, Adelson K, Hoshida Y, Koh AP, Shah N, Mandeli J, Ting J, Germain D. Patient-derived interstitial fluids and predisposition to aggressive sporadic breast cancer through collagen remodeling and inactivation of p53. Clin Cancer Res. 2017;23:5446–59.ArticleCASPubMedPubMed CentralGoogle Scholar
Wormann SM, Song L, Ai J, Diakopoulos KN, Kurkowski MU, Gorgulu K, Ruess D, Campbell A, Doglioni C, Jodrell D, et al. Loss of P53 function activates JAK2-STAT3 signaling to promote pancreatic tumor growth, stroma modification, and gemcitabine resistance in mice and is associated with patient survival. Gastroenterology. 2016;151(180–193):e112.Google Scholar
Teodoro JG, Parker AE, Zhu X, Green MR. p53-mediated inhibition of angiogenesis through up-regulation of a collagen prolyl hydroxylase. Science. 2006;313:968–71.ArticleCASPubMedGoogle Scholar
Assadian S, El-Assaad W, Wang XQ, Gannon PO, Barres V, Latour M, Mes-Masson AM, Saad F, Sado Y, Dostie J, Teodoro JG. p53 inhibits angiogenesis by inducing the production of Arresten. Cancer Res. 2012;72:1270–9.ArticleCASPubMedGoogle Scholar
Jolly LA, Novitskiy S, Owens P, Massoll N, Cheng N, Fang W, Moses HL, Franco AT. Fibroblast-mediated collagen remodeling within the tumor microenvironment facilitates progression of thyroid cancers driven by BrafV600E and Pten loss. Cancer Res. 2016;76:1804–13.ArticleCASPubMedPubMed CentralGoogle Scholar
Shields MA, Ebine K, Sahai V, Kumar K, Siddiqui K, Hwang RF, Grippo PJ, Munshi HG. Snail cooperates with KrasG12D to promote pancreatic fibrosis. Mol Cancer Res. 2013;11:1078–87.ArticleCASPubMedPubMed CentralGoogle Scholar
Wang JH, Newbury LJ, Knisely AS, Monia B, Hendry BM, Sharpe CC. Antisense knockdown of Kras inhibits fibrosis in a rat model of unilateral ureteric obstruction. Am J Pathol. 2012;180:82–90.ArticleCASPubMedGoogle Scholar
Yoshida T, Hashimura M, Kuwata T, Matsumoto T, Suzuki E, Tazo Y, Nakajima H, Inukai M, Saegusa M. Transcriptional regulation of the alpha-1 type II collagen gene by nuclear factor B/p65 and Sox9 in the chondrocytic phenotype of uterine carcinosarcomas. Hum Pathol. 2013;44:1780–8.ArticleCASPubMedGoogle Scholar
Nagathihalli NS, Castellanos JA, Shi C, Beesetty Y, Reyzer ML, Caprioli R, Chen X, Walsh AJ, Skala MC, Moses HL, Merchant NB. Signal transducer and activator of transcription 3, mediated remodeling of the tumor microenvironment results in enhanced tumor drug delivery in a mouse model of pancreatic cancer. Gastroenterology. 2015;149(1932–1943):e1939.Google Scholar
Laklai H, Miroshnikova YA, Pickup MW, Collisson EA, Kim GE, Barrett AS, Hill RC, Lakins JN, Schlaepfer DD, Mouw JK, et al. Genotype tunes pancreatic ductal adenocarcinoma tissue tension to induce matricellular fibrosis and tumor progression. Nat Med. 2016;22:497–505.ArticleCASPubMedPubMed CentralGoogle Scholar
Miskolczi Z, Smith MP, Rowling EJ, Ferguson J, Barriuso J, Wellbrock C. Collagen abundance controls melanoma phenotypes through lineage-specific microenvironment sensing. Oncogene. 2018;37:3166–82.ArticleCASPubMedPubMed CentralGoogle Scholar
Jenkins MH, Croteau W, Mullins DW, Brinckerhoff CE. The BRAF(V600E) inhibitor, PLX4032, increases type I collagen synthesis in melanoma cells. Matrix Biol. 2015;48:66–77.ArticleCASPubMedPubMed CentralGoogle Scholar
Clarke CJ, Berg TJ, Birch J, Ennis D, Mitchell L, Cloix C, Campbell A, Sumpton D, Nixon C, Campbell K, et al. The initiator methionine tRNA drives secretion of type II collagen from stromal fibroblasts to promote tumor growth and angiogenesis. Curr Biol. 2016;26:755–65.ArticleCASPubMedPubMed CentralGoogle Scholar
Sugiyama N, Varjosalo M, Meller P, Lohi J, Hyytiainen M, Kilpinen S, Kallioniemi O, Ingvarsen S, Engelholm LH, Taipale J, et al. Fibroblast growth factor receptor 4 regulates tumor invasion by coupling fibroblast growth factor signaling to extracellular matrix degradation. Cancer Res. 2010;70:7851–61.ArticleCASPubMedGoogle Scholar
Holdman XB, Welte T, Rajapakshe K, Pond A, Coarfa C, Mo Q, Huang S, Hilsenbeck SG, Edwards DP, Zhang X, Rosen JM. Upregulation of EGFR signaling is correlated with tumor stroma remodeling and tumor recurrence in FGFR1-driven breast cancer. Breast Cancer Res. 2015;17:141.ArticlePubMedPubMed CentralCASGoogle Scholar
Chang CC, Hsieh TL, Tiong TY, Hsiao CH, Ji AT, Hsu WT, Lee OK, Ho JH. Regulation of metastatic ability and drug resistance in pulmonary adenocarcinoma by matrix rigidity via activating c-Met and EGFR. Biomaterials. 2015;60:141–50.ArticleCASPubMedGoogle Scholar
Wei SC, Fattet L, Tsai JH, Guo Y, Pai VH, Majeski HE, Chen AC, Sah RL, Taylor SS, Engler AJ, Yang J. Matrix stiffness drives epithelial-mesenchymal transition and tumour metastasis through a TWIST1-G3BP2 mechanotransduction pathway. Nat Cell Biol. 2015;17:678–88.ArticleCASPubMedPubMed CentralGoogle Scholar
Sipes NS, Feng Y, Guo F, Lee HO, Chou FS, Cheng J, Mulloy J, Zheng Y. Cdc42 regulates extracellular matrix remodeling in three dimensions. J Biol Chem. 2011;286:36469–77.ArticleCASPubMedPubMed CentralGoogle Scholar
Yamazaki D, Kurisu S, Takenawa T. Involvement of Rac and Rho signaling in cancer cell motility in 3D substrates. Oncogene. 2009;28:1570–83.ArticleCASPubMedGoogle Scholar
Meng C, He Y, Wei Z, Lu Y, Du F, Ou G, Wang N, Luo XG, Ma W, Zhang TC, He H. MRTF-A mediates the activation of COL1A1 expression stimulated by multiple signaling pathways in human breast cancer cells. Biomed Pharmacother. 2018;104:718–28.ArticleCASPubMedGoogle Scholar
Vennin C, Chin VT, Warren SC, Lucas MC, Herrmann D, Magenau A, Melenec P, Walters SN, Del Monte-Nieto G, Conway JR, et al. Transient tissue priming via ROCK inhibition uncouples pancreatic cancer progression, sensitivity to chemotherapy, and metastasis. Sci Transl Med. 2017. https://doi.org/10.1126/scitranslmed.aai8504.ArticlePubMedPubMed CentralGoogle Scholar
Rath N, Morton JP, Julian L, Helbig L, Kadir S, McGhee EJ, Anderson KI, Kalna G, Mullin M, Pinho AV, et al. ROCK signaling promotes collagen remodeling to facilitate invasive pancreatic ductal adenocarcinoma tumor cell growth. EMBO Mol Med. 2017;9:198–218.ArticleCASPubMedGoogle Scholar
Favreau AJ, Vary CP, Brooks PC, Sathyanarayana P. Cryptic collagen IV promotes cell migration and adhesion in myeloid leukemia. Cancer Med. 2014;3:265–72.ArticleCASPubMedPubMed CentralGoogle Scholar
Huang H, Svoboda RA, Lazenby AJ, Saowapa J, Chaika N, Ding K, Wheelock MJ, Johnson KR. Up-regulation of N-cadherin by Collagen I-activated discoidin domain receptor 1 in pancreatic cancer requires the adaptor molecule Shc1. J Biol Chem. 2016;291:23208–23.ArticleCASPubMedPubMed CentralGoogle Scholar
Zhang K, Corsa CA, Ponik SM, Prior JL, Piwnica-Worms D, Eliceiri KW, Keely PJ, Longmore GD. The collagen receptor discoidin domain receptor 2 stabilizes SNAIL1 to facilitate breast cancer metastasis. Nat Cell Biol. 2013;15:677–87.ArticleCASPubMedPubMed CentralGoogle Scholar
Iwai LK, Payne LS, Luczynski MT, Chang F, Xu H, Clinton RW, Paul A, Esposito EA, Gridley S, Leitinger B, et al. Phosphoproteomics of collagen receptor networks reveals SHP-2 phosphorylation downstream of wild-type DDR2 and its lung cancer mutants. Biochem J. 2013;454:501–13.ArticleCASPubMedGoogle Scholar
Rada M, Nallanthighal S, Cha J, Ryan K, Sage J, Eldred C, Ullo M, Orsulic S, Cheon DJ. Inhibitor of apoptosis proteins (IAPs) mediate collagen type XI alpha 1-driven cisplatin resistance in ovarian cancer. Oncogene. 2018;37:4809–20.ArticleCASPubMedGoogle Scholar
Oliveira-Ferrer L, Rossler K, Haustein V, Schroder C, Wicklein D, Maltseva D, Khaustova N, Samatov T, Tonevitsky A, Mahner S, et al. c-FOS suppresses ovarian cancer progression by changing adhesion. Br J Cancer. 2014;110:753–63.ArticleCASPubMedGoogle Scholar
Procacci P, Moscheni C, Sartori P, Sommariva M, Gagliano N. Tumor(-)stroma cross-talk in human pancreatic ductal adenocarcinoma: a focus on the effect of the extracellular matrix on tumor cell phenotype and invasive potential. Cells. 2018;7:E158.ArticlePubMedGoogle Scholar
Imamichi Y, Konig A, Gress T, Menke A. Collagen type I-induced Smad-interacting protein 1 expression downregulates E-cadherin in pancreatic cancer. Oncogene. 2007;26:2381–5.ArticleCASPubMedGoogle Scholar
Vaniotis G, Rayes RF, Qi S, Milette S, Wang N, Perrino S, Bourdeau F, Nystrom H, He Y, Lamarche-Vane N, Brodt P. Collagen IV-conveyed signals can regulate chemokine production and promote liver metastasis. Oncogene. 2018;37:3790–805.ArticleCASPubMedGoogle Scholar
Espinosa Neira R, Salazar EP. Native type IV collagen induces an epithelial to mesenchymal transition-like process in mammary epithelial cells MCF10A. Int J Biochem Cell Biol. 2012;44:2194–203.ArticleCASPubMedGoogle Scholar
Spivey KA, Chung I, Banyard J, Adini I, Feldman HA, Zetter BR. A role for collagen XXIII in cancer cell adhesion, anchorage-independence and metastasis. Oncogene. 2012;31:2362–72.ArticleCASPubMedGoogle Scholar
Hayashido Y, Kitano H, Sakaue T, Fujii T, Suematsu M, Sakurai S, Okamoto T. Overexpression of integrin alphav facilitates proliferation and invasion of oral squamous cell carcinoma cells via MEK/ERK signaling pathway that is activated by interaction of integrin alphavbeta8 with type collagen. Int J Oncol. 2014;45:1875–82.ArticleCASPubMedGoogle Scholar
Ibbetson SJ, Pyne NT, Pollard AN, Olson MF, Samuel MS. Mechanotransduction pathways promoting tumor progression are activated in invasive human squamous cell carcinoma. Am J Pathol. 2013;183:930–7.ArticleCASPubMedGoogle Scholar
Shen Y, Shen R, Ge L, Zhu Q, Li F. Fibrillar type I collagen matrices enhance metastasis/invasion of ovarian epithelial cancer via beta1 integrin and PTEN signals. Int J Gynecol Cancer. 2012;22:1316–24.ArticlePubMedGoogle Scholar
Girotti MR, Fernandez M, Lopez JA, Camafeita E, Fernandez EA, Albar JP, Benedetti LG, Valacco MP, Brekken RA, Podhajcer OL, Llera AS. SPARC promotes cathepsin B-mediated melanoma invasiveness through a collagen I/alpha2beta1 integrin axis. J Invest Dermatol. 2011;131:2438–47.ArticleCASPubMedGoogle Scholar
Kirkland SC. Type I collagen inhibits differentiation and promotes a stem cell-like phenotype in human colorectal carcinoma cells. Br J Cancer. 2009;101:320–6.ArticleCASPubMedPubMed CentralGoogle Scholar
Smith SD, Enge M, Bao W, Thullberg M, Costa TD, Olofsson H, Gashi B, Selivanova G, Stromblad S. Protein kinase Calpha (PKCalpha) regulates p53 localization and melanoma cell survival downstream of integrin alphav in three-dimensional collagen and in vivo. J Biol Chem. 2012;287:29336–47.ArticleCASPubMedPubMed CentralGoogle Scholar
Hwangbo C, Park J, Lee JH. mda-9/Syntenin protein positively regulates the activation of Akt protein by facilitating integrin-linked kinase adaptor function during adhesion to type I collagen. J Biol Chem. 2011;286:33601–12.ArticleCASPubMedPubMed CentralGoogle Scholar
Blockhuys S, Van Rompaye B, De Rycke R, Lambein K, Claes K, Bracke M, De Wagter C, De Wever O. Radiation-induced myosin IIA expression stimulates collagen type I matrix reorganization. Radiother Oncol. 2013;108:162–7.ArticleCASPubMedGoogle Scholar
Artym VV, Swatkoski S, Matsumoto K, Campbell CB, Petrie RJ, Dimitriadis EK, Li X, Mueller SC, Bugge TH, Gucek M, Yamada KM. Dense fibrillar collagen is a potent inducer of invadopodia via a specific signaling network. J Cell Biol. 2015;208:331–50.ArticlePubMedPubMed CentralGoogle Scholar
Yan T, Zhang A, Shi F, Chang F, Mei J, Liu Y, Zhu Y. Integrin alphavbeta3-associated DAAM1 is essential for collagen-induced invadopodia extension and cell haptotaxis in breast cancer cells. J Biol Chem. 2018;293:10172–85.ArticleCASPubMedPubMed CentralGoogle Scholar
Chen SY, Lin JS, Yang BC. Modulation of tumor cell stiffness and migration by type IV collagen through direct activation of integrin signaling pathway. Arch Biochem Biophys. 2014;555–556:1–8.ArticlePubMedCASGoogle Scholar
Hodgson L, Henderson AJ, Dong C. Melanoma cell migration to type IV collagen requires activation of NF-kappaB. Oncogene. 2003;22:98–108.ArticleCASPubMedPubMed CentralGoogle Scholar
Cattaruzza S, Nicolosi PA, Braghetta P, Pazzaglia L, Benassi MS, Picci P, Lacrima K, Zanocco D, Rizzo E, Stallcup WB, et al. NG2/CSPG4-collagen type VI interplays putatively involved in the microenvironmental control of tumour engraftment and local expansion. J Mol Cell Biol. 2013;5:176–93.ArticleCASPubMedPubMed CentralGoogle Scholar
Zhang H, Fredericks T, Xiong G, Qi Y, Rychahou PG, Li JD, Pihlajaniemi T, Xu W, Xu R. Membrane associated collagen XIII promotes cancer metastasis and enhances anoikis resistance. Breast Cancer Res. 2018;20:116.ArticlePubMedPubMed CentralCASGoogle Scholar
Bauer R, Ratzinger S, Wales L, Bosserhoff A, Senner V, Grifka J, Grassel S. Inhibition of collagen XVI expression reduces glioma cell invasiveness. Cell Physiol Biochem. 2011;27:217–26.ArticleCASPubMedGoogle Scholar
Ratzinger S, Grassel S, Dowejko A, Reichert TE, Bauer RJ. Induction of type XVI collagen expression facilitates proliferation of oral cancer cells. Matrix Biol. 2011;30:118–25.ArticleCASPubMedGoogle Scholar
Stawikowski MJ, Aukszi B, Stawikowska R, Cudic M, Fields GB. Glycosylation modulates melanoma cell alpha2beta1 and alpha3beta1 integrin interactions with type IV collagen. J Biol Chem. 2014;289:21591–604.ArticlePubMedPubMed CentralCASGoogle Scholar
Liu S, Liao G, Li G. Regulatory effects of COL1A1 on apoptosis induced by radiation in cervical cancer cells. Cancer Cell Int. 2017;17:73.ArticlePubMedPubMed CentralCASGoogle Scholar
Shea MP, O’Leary KA, Wegner KA, Vezina CM, Schuler LA. High collagen density augments mTOR-dependent cancer stem cells in ERalpha + mammary carcinomas, and increases mTOR-independent lung metastases. Cancer Lett. 2018;433:1–9.ArticleCASPubMedPubMed CentralGoogle Scholar
Barcus CE, O’Leary KA, Brockman JL, Rugowski DE, Liu Y, Garcia N, Yu M, Keely PJ, Eliceiri KW, Schuler LA. Elevated collagen-I augments tumor progressive signals, intravasation and metastasis of prolactin-induced estrogen receptor alpha positive mammary tumor cells. Breast Cancer Res. 2017;19:9.ArticlePubMedPubMed CentralCASGoogle Scholar
Yamazaki S, Higuchi Y, Ishibashi M, Hashimoto H, Yasunaga M, Matsumura Y, Tsuchihara K, Tsuboi M, Goto K, Ochiai A, Ishii G. Collagen type I induces EGFR-TKI resistance in EGFR-mutated cancer cells by mTOR activation through Akt-independent pathway. Cancer Sci. 2018;109:2063–73.ArticleCASPubMedPubMed CentralGoogle Scholar
Brown CW, Brodsky AS, Freiman RN. Notch3 overexpression promotes anoikis resistance in epithelial ovarian cancer via upregulation of COL4A2. Mol Cancer Res. 2015;13:78–85.ArticleCASPubMedGoogle Scholar
Said G, Guilbert M, Millerot-Serrurot E, Van Gulick L, Terryn C, Garnotel R, Jeannesson P. Impact of carbamylation and glycation of collagen type I on migration of HT1080 human fibrosarcoma cells. Int J Oncol. 2012;40:1797–804.CASPubMedGoogle Scholar
Liu J, Kang SG, Wang P, Wang Y, Lv X, Liu Y, Wang F, Gu Z, Yang Z, Weber JK, et al. Molecular mechanism of Gd@C82(OH)22 increasing collagen expression: implication for encaging tumor. Biomaterials. 2018;152:24–36.ArticleCASPubMedGoogle Scholar
Fang T, Lv H, Lv G, Li T, Wang C, Han Q, Yu L, Su B, Guo L, Huang S, et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat Commun. 2018;9:191.ArticlePubMedPubMed CentralCASGoogle Scholar
Liu X, Cao M, Palomares M, Wu X, Li A, Yan W, Fong MY, Chan WC, Wang SE. Metastatic breast cancer cells overexpress and secrete miR-218 to regulate type I collagen deposition by osteoblasts. Breast Cancer Res. 2018;20:127.ArticleCASPubMedPubMed CentralGoogle Scholar
Colden M, Dar AA, Saini S, Dahiya PV, Shahryari V, Yamamura S, Tanaka Y, Stein G, Dahiya R, Majid S. MicroRNA-466 inhibits tumor growth and bone metastasis in prostate cancer by direct regulation of osteogenic transcription factor RUNX2. Cell Death Dis. 2017;8:e2572.ArticleCASPubMedPubMed CentralGoogle Scholar
Li AL, Chung TS, Chan YN, Chen CL, Lin SC, Chiang YR, Lin CH, Chen CC, Ma N. microRNA expression pattern as an ancillary prognostic signature for radiotherapy. J Transl Med. 2018;16:341.ArticleCASPubMedPubMed CentralGoogle Scholar
Rabieian R, Boshtam M, Zareei M, Kouhpayeh S, Masoudifar A, Mirzaei H. Plasminogen activator inhibitor type-1 as a regulator of fibrosis. J Cell Biochem. 2018;119:17–27.ArticleCASPubMedGoogle Scholar
Mirzaei H, Ferns GA, Avan A, Mobarhan MG. Cytokines and MicroRNA in coronary artery disease. Adv Clin Chem. 2017;82:47–70.ArticleCASPubMedGoogle Scholar
Simonian M, Mosallayi M, Mirzaei H. Circulating miR-21 as novel biomarker in gastric cancer: diagnostic and prognostic biomarker. J Cancer Res Ther. 2018;14:475.PubMedGoogle Scholar
Saeedi Borujeni MJ, Esfandiary E, Taheripak G, Codoner-Franch P, Alonso-Iglesias E, Mirzaei H. Molecular aspects of diabetes mellitus: resistin, microRNA, and exosome. J Cell Biochem. 2018;119:1257–72.ArticleCASPubMedGoogle Scholar
Hoey C, Ahmed M, Fotouhi Ghiam A, Vesprini D, Huang X, Commisso K, Commisso A, Ray J, Fokas E, Loblaw DA, et al. Circulating miRNAs as non-invasive biomarkers to predict aggressive prostate cancer after radical prostatectomy. J Transl Med. 2019;17:173.ArticleCASPubMedPubMed CentralGoogle Scholar
Lin Y, Chen F, Shen L, Tang X, Du C, Sun Z, Ding H, Chen J, Shen B. Biomarker microRNAs for prostate cancer metastasis: screened with a network vulnerability analysis model. J Transl Med. 2018;16:134.ArticleCASPubMedPubMed CentralGoogle Scholar
Keshavarzi M, Sorayayi S, Jafar Rezaei M, Mohammadi M, Ghaderi A, Rostamzadeh A, Masoudifar A, Mirzaei H. MicroRNAs-based imaging techniques in cancer diagnosis and therapy. J Cell Biochem. 2017;118:4121–8.ArticleCASPubMedGoogle Scholar
Gholamin S, Mirzaei H, Razavi SM, Hassanian SM, Saadatpour L, Masoudifar A, ShahidSales S, Avan A. GD2-targeted immunotherapy and potential value of circulating microRNAs in neuroblastoma. J Cell Physiol. 2018;233:866–79.ArticleCASPubMedGoogle Scholar
Jamali L, Tofigh R, Tutunchi S, Panahi G, Borhani F, Akhavan S, Nourmohammadi P, Ghaderian SMH, Rasouli M, Mirzaei H. Circulating microRNAs as diagnostic and therapeutic biomarkers in gastric and esophageal cancers. J Cell Physiol. 2018;233:8538–50.ArticleCASPubMedGoogle Scholar
Zhu X, Rao X, Yao W, Zou X. Downregulation of MiR-196b-5p impedes cell proliferation and metastasis in breast cancer through regulating COL1A1. Am J Transl Res. 2018;10:3122–32.PubMedPubMed CentralGoogle Scholar
Lai YH, Chen J, Wang XP, Wu YQ, Peng HT, Lin XH, Wang WJ. Collagen triple helix repeat containing-1 negatively regulated by microRNA-30c promotes cell proliferation and metastasis and indicates poor prognosis in breast cancer. J Exp Clin Cancer Res. 2017;36:92.ArticlePubMedPubMed CentralCASGoogle Scholar
La T, Liu GZ, Farrelly M, Cole N, Feng YC, Zhang YY, Sherwin SK, Yari H, Tabatabaee H, Yan XG, et al. A p53-responsive miRNA network promotes cancer cell quiescence. Cancer Res. 2018;78:6666–79.ArticleCASPubMedGoogle Scholar
Asama H, Suzuki R, Hikichi T, Takagi T, Masamune A, Ohira H. MicroRNA let-7d targets thrombospondin-1 and inhibits the activation of human pancreatic stellate cells. Pancreatology. 2019;19:196–203.ArticleCASPubMedGoogle Scholar
Su B, Zhao W, Shi B, Zhang Z, Yu X, Xie F, Guo Z, Zhang X, Liu J, Shen Q, et al. Let-7d suppresses growth, metastasis, and tumor macrophage infiltration in renal cell carcinoma by targeting COL3A1 and CCL7. Mol Cancer. 2014;13:206.ArticlePubMedPubMed CentralCASGoogle Scholar
Yamamoto N, Kinoshita T, Nohata N, Yoshino H, Itesako T, Fujimura L, Mitsuhashi A, Usui H, Enokida H, Nakagawa M, et al. Tumor-suppressive microRNA-29a inhibits cancer cell migration and invasion via targeting HSP47 in cervical squamous cell carcinoma. Int J Oncol. 2013;43:1855–63.ArticleCASPubMedPubMed CentralGoogle Scholar
Zhao Y, Cao J, Melamed A, Worley M, Gockley A, Jones D, Nia HT, Zhang Y, Stylianopoulos T, Kumar AS, et al. Losartan treatment enhances chemotherapy efficacy and reduces ascites in ovarian cancer models by normalizing the tumor stroma. Proc Natl Acad Sci USA. 2019;116:2210–9.ArticlePubMedCASPubMed CentralGoogle Scholar
Ramanathan S, Douglas SR, Alexander GM, Shenoda BB, Barrett JE, Aradillas E, Sacan A, Ajit SK. Exosome microRNA signatures in patients with complex regional pain syndrome undergoing plasma exchange. J Transl Med. 2019;17:81.ArticlePubMedPubMed CentralGoogle Scholar
Yeon JH, Jeong HE, Seo H, Cho S, Kim K, Na D, Chung S, Park J, Choi N, Kang JY. Cancer-derived exosomes trigger endothelial to mesenchymal transition followed by the induction of cancer-associated fibroblasts. Acta Biomater. 2018;76:146–53.ArticleCASPubMedGoogle Scholar
Hakulinen J, Sankkila L, Sugiyama N, Lehti K, Keski-Oja J. Secretion of active membrane type 1 matrix metalloproteinase (MMP-14) into extracellular space in microvesicular exosomes. J Cell Biochem. 2008;105:1211–8.ArticleCASPubMedGoogle Scholar
Szvicsek Z, Oszvald A, Szabo L, Sandor GO, Kelemen A, Soos AA, Paloczi K, Harsanyi L, Tolgyes T, Dede K, et al. Extracellular vesicle release from intestinal organoids is modulated by Apc mutation and other colorectal cancer progression factors. Cell Mol Life Sci. 2019;76:2463–76.ArticleCASPubMedPubMed CentralGoogle Scholar
Kakkad SM, Solaiyappan M, O’Rourke B, Stasinopoulos I, Ackerstaff E, Raman V, Bhujwalla ZM, Glunde K. Hypoxic tumor microenvironments reduce collagen I fiber density. Neoplasia. 2010;12:608–17.ArticleCASPubMedPubMed CentralGoogle Scholar
Di Stefano V, Torsello B, Bianchi C, Cifola I, Mangano E, Bovo G, Cassina V, De Marco S, Corti R, Meregalli C, et al. Major action of endogenous lysyl Oxidase in clear cell renal cell carcinoma progression and collagen stiffness revealed by primary cell cultures. Am J Pathol. 2016;186:2473–85.ArticlePubMedCASGoogle Scholar
Wong CC, Gilkes DM, Zhang H, Chen J, Wei H, Chaturvedi P, Fraley SI, Wong CM, Khoo US, Ng IO, et al. Hypoxia-inducible factor 1 is a master regulator of breast cancer metastatic niche formation. Proc Natl Acad Sci USA. 2011;108:16369–74.ArticleCASPubMedPubMed CentralGoogle Scholar
Tse AP, Sze KM, Shea QT, Chiu EY, Tsang FH, Chiu DK, Zhang MS, Lee D, Xu IM, Chan CY, et al. Hepatitis transactivator protein X promotes extracellular matrix modification through HIF/LOX pathway in liver cancer. Oncogenesis. 2018;7:44.ArticlePubMedPubMed CentralCASGoogle Scholar
Gao Y, Xiao Q, Ma H, Li L, Liu J, Feng Y, Fang Z, Wu J, Han X, Zhang J, et al. LKB1 inhibits lung cancer progression through lysyl oxidase and extracellular matrix remodeling. Proc Natl Acad Sci USA. 2010;107:18892–7.ArticleCASPubMedPubMed CentralGoogle Scholar
Eisinger-Mathason TS, Zhang M, Qiu Q, Skuli N, Nakazawa MS, Karakasheva T, Mucaj V, Shay JE, Stangenberg L, Sadri N, et al. Hypoxia-dependent modification of collagen networks promotes sarcoma metastasis. Cancer Discov. 2013;3:1190–205.ArticleCASPubMedGoogle Scholar
Lewis DM, Pruitt H, Jain N, Ciccaglione M, McCaffery JM, Xia Z, Weber K, Eisinger-Mathason TSK, Gerecht S. A feedback loop between hypoxia and matrix stress relaxation increases oxygen-axis migration and metastasis in sarcoma. Cancer Res. 2019;79:1981–95.ArticleCASPubMedPubMed CentralGoogle Scholar
Goggins E, Kakkad S, Mironchik Y, Jacob D, Wildes F, Krishnamachary B, Bhujwalla ZM. Hypoxia inducible factors modify collagen I fibers in MDA-MB-231 triple negative breast cancer xenografts. Neoplasia. 2018;20:131–9.ArticleCASPubMedGoogle Scholar
Xiong G, Stewart RL, Chen J, Gao T, Scott TL, Samayoa LM, O’Connor K, Lane AN, Xu R. Collagen prolyl 4-hydroxylase 1 is essential for HIF-1alpha stabilization and TNBC chemoresistance. Nat Commun. 2018;9:4456.ArticlePubMedPubMed CentralCASGoogle Scholar
Amelio I, Mancini M, Petrova V, Cairns RA, Vikhreva P, Nicolai S, Marini A, Antonov AA, Le Quesne J, Baena Acevedo JD, et al. p53 mutants cooperate with HIF-1 in transcriptional regulation of extracellular matrix components to promote tumor progression. Proc Natl Acad Sci USA. 2018;115:E10869–78.ArticleCASPubMedPubMed CentralGoogle Scholar
Whittington CF, Yoder MC, Voytik-Harbin SL. Collagen-polymer guidance of vessel network formation and stabilization by endothelial colony forming cells in vitro. Macromol Biosci. 2013;13:1135–49.ArticleCASPubMedGoogle Scholar
Bordeleau F, Mason BN, Lollis EM, Mazzola M, Zanotelli MR, Somasegar S, Califano JP, Montague C, LaValley DJ, Huynh J, et al. Matrix stiffening promotes a tumor vasculature phenotype. Proc Natl Acad Sci USA. 2017;114:492–7.ArticleCASPubMedGoogle Scholar
Kurban G, Duplan E, Ramlal N, Hudon V, Sado Y, Ninomiya Y, Pause A. Collagen matrix assembly is driven by the interaction of von Hippel-Lindau tumor suppressor protein with hydroxylated collagen IV alpha 2. Oncogene. 2008;27:1004–12.ArticleCASPubMedGoogle Scholar
Burmakin M, van Wieringen T, Olsson PO, Stuhr L, Ahgren A, Heldin CH, Reed RK, Rubin K, Hellberg C. Imatinib increases oxygen delivery in extracellular matrix-rich but not in matrix-poor experimental carcinoma. J Transl Med. 2017;15:47.ArticlePubMedPubMed CentralCASGoogle Scholar
Mahajan UM, Langhoff E, Goni E, Costello E, Greenhalf W, Halloran C, Ormanns S, Kruger S, Boeck S, Ribback S, et al. Immune cell and stromal signature associated with progression-free survival of patients with resected pancreatic ductal adenocarcinoma. Gastroenterology. 2018;155(1625–1639):e1622.Google Scholar
Batista ML Jr, Henriques FS, Neves RX, Olivan MR, Matos-Neto EM, Alcantara PS, Maximiano LF, Otoch JP, Alves MJ, Seelaender M. Cachexia-associated adipose tissue morphological rearrangement in gastrointestinal cancer patients. J Cachexia Sarcopenia Muscle. 2016;7:37–47.ArticlePubMedGoogle Scholar
Brodsky AS, Xiong J, Yang D, Schorl C, Fenton MA, Graves TA, Sikov WM, Resnick MB, Wang Y. Identification of stromal ColXalpha1 and tumor-infiltrating lymphocytes as putative predictive markers of neoadjuvant therapy in estrogen receptor-positive/HER2-positive breast cancer. BMC Cancer. 2016;16:274.ArticlePubMedPubMed CentralCASGoogle Scholar
Kuczek DE, Larsen AMH, Thorseth ML, Carretta M, Kalvisa A, Siersbaek MS, Simoes AMC, Roslind A, Engelholm LH, Noessner E, et al. Collagen density regulates the activity of tumor-infiltrating T cells. J Immunother Cancer. 2019;7:68.ArticlePubMedPubMed CentralGoogle Scholar
Bougherara H, Mansuet-Lupo A, Alifano M, Ngo C, Damotte D, Le Frere-Belda MA, Donnadieu E, Peranzoni E. Real-time imaging of resident t cells in human lung and ovarian carcinomas reveals how different tumor microenvironments control T lymphocyte migration. Front Immunol. 2015;6:500.ArticlePubMedPubMed CentralGoogle Scholar
Salmon H, Franciszkiewicz K, Damotte D, Dieu-Nosjean MC, Validire P, Trautmann A, Mami-Chouaib F, Donnadieu E. Matrix architecture defines the preferential localization and migration of T cells into the stroma of human lung tumors. J Clin Invest. 2012;122:899–910.ArticleCASPubMedPubMed CentralGoogle Scholar
Carstens JL, Correa de Sampaio P, Yang D, Barua S, Wang H, Rao A, Allison JP, LeBleu VS, Kalluri R. Spatial computation of intratumoral T cells correlates with survival of patients with pancreatic cancer. Nat Commun. 2017;8:15095.ArticlePubMedPubMed CentralGoogle Scholar
Mariathasan S, Turley SJ, Nickles D, Castiglioni A, Yuen K, Wang Y, Kadel EE III, Koeppen H, Astarita JL, Cubas R, et al. TGFbeta attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554:544–8.ArticleCASPubMedPubMed CentralGoogle Scholar
Budhu S, Schaer DA, Li Y, Toledo-Crow R, Panageas K, Yang X, Zhong H, Houghton AN, Silverstein SC, Merghoub T, Wolchok JD. Blockade of surface-bound TGF-beta on regulatory T cells abrogates suppression of effector T cell function in the tumor microenvironment. Sci Signal. 2017;10:eaak9702.ArticlePubMedPubMed CentralCASGoogle Scholar
Pinho AV, Van Bulck M, Chantrill L, Arshi M, Sklyarova T, Herrmann D, Vennin C, Gallego-Ortega D, Mawson A, Giry-Laterriere M, et al. ROBO2 is a stroma suppressor gene in the pancreas and acts via TGF-beta signalling. Nat Commun. 2018;9:5083.ArticlePubMedPubMed CentralCASGoogle Scholar
Acerbi I, Cassereau L, Dean I, Shi Q, Au A, Park C, Chen YY, Liphardt J, Hwang ES, Weaver VM. Human breast cancer invasion and aggression correlates with ECM stiffening and immune cell infiltration. Integr Biol (Camb). 2015;7:1120–34.ArticleCASGoogle Scholar
O’Brien J, Lyons T, Monks J, Lucia MS, Wilson RS, Hines L, Man YG, Borges V, Schedin P. Alternatively activated macrophages and collagen remodeling characterize the postpartum involuting mammary gland across species. Am J Pathol. 2010;176:1241–55.ArticlePubMedPubMed CentralCASGoogle Scholar
Shi C, Washington MK, Chaturvedi R, Drosos Y, Revetta FL, Weaver CJ, Buzhardt E, Yull FE, Blackwell TS, Sosa-Pineda B, et al. Fibrogenesis in pancreatic cancer is a dynamic process regulated by macrophage-stellate cell interaction. Lab Invest. 2014;94:409–21.ArticleCASPubMedPubMed CentralGoogle Scholar
Afik R, Zigmond E, Vugman M, Klepfish M, Shimshoni E, Pasmanik-Chor M, Shenoy A, Bassat E, Halpern Z, Geiger T, et al. Tumor macrophages are pivotal constructors of tumor collagenous matrix. J Exp Med. 2016;213:2315–31.ArticleCASPubMedPubMed CentralGoogle Scholar
Madsen DH, Ingvarsen S, Jurgensen HJ, Melander MC, Kjoller L, Moyer A, Honore C, Madsen CA, Garred P, Burgdorf S, et al. The non-phagocytic route of collagen uptake: a distinct degradation pathway. J Biol Chem. 2011;286:26996–7010.ArticleCASPubMedPubMed CentralGoogle Scholar
Madsen DH, Leonard D, Masedunskas A, Moyer A, Jurgensen HJ, Peters DE, Amornphimoltham P, Selvaraj A, Yamada SS, Brenner DA, et al. M2-like macrophages are responsible for collagen degradation through a mannose receptor-mediated pathway. J Cell Biol. 2013;202:951–66.ArticleCASPubMedPubMed CentralGoogle Scholar
Madsen DH, Jurgensen HJ, Siersbaek MS, Kuczek DE, Grey Cloud L, Liu S, Behrendt N, Grontved L, Weigert R, Bugge TH. Tumor-associated macrophages derived from circulating inflammatory monocytes degrade collagen through cellular uptake. Cell Rep. 2017;21:3662–71.ArticleCASPubMedPubMed CentralGoogle Scholar
Muliaditan T, Caron J, Okesola M, Opzoomer JW, Kosti P, Georgouli M, Gordon P, Lall S, Kuzeva DM, Pedro L, et al. Macrophages are exploited from an innate wound healing response to facilitate cancer metastasis. Nat Commun. 2018;9:2951.ArticlePubMedPubMed CentralCASGoogle Scholar
Esbona K, Inman D, Saha S, Jeffery J, Schedin P, Wilke L, Keely P. COX-2 modulates mammary tumor progression in response to collagen density. Breast Cancer Res. 2016;18:35.ArticlePubMedPubMed CentralCASGoogle Scholar
Cui X, Morales RT, Qian W, Wang H, Gagner JP, Dolgalev I, Placantonakis D, Zagzag D, Cimmino L, Snuderl M, et al. Hacking macrophage-associated immunosuppression for regulating glioblastoma angiogenesis. Biomaterials. 2018;161:164–78.ArticleCASPubMedPubMed CentralGoogle Scholar
Tevis KM, Cecchi RJ, Colson YL, Grinstaff MW. Mimicking the tumor microenvironment to regulate macrophage phenotype and assessing chemotherapeutic efficacy in embedded cancer cell/macrophage spheroid models. Acta Biomater. 2017;50:271–9.ArticleCASPubMedGoogle Scholar
Ma PF, Gao CC, Yi J, Zhao JL, Liang SQ, Zhao Y, Ye YC, Bai J, Zheng QJ, Dou KF, et al. Cytotherapy with M1-polarized macrophages ameliorates liver fibrosis by modulating immune microenvironment in mice. J Hepatol. 2017;67:770–9.ArticleCASPubMedGoogle Scholar
Staser K, Yang FC, Clapp DW. Pathogenesis of plexiform neurofibroma: tumor-stromal/hematopoietic interactions in tumor progression. Annu Rev Pathol. 2012;7:469–95.ArticleCASPubMedGoogle Scholar
Garcia-Mendoza MG, Inman DR, Ponik SM, Jeffery JJ, Sheerar DS, Van Doorn RR, Keely PJ. Neutrophils drive accelerated tumor progression in the collagen-dense mammary tumor microenvironment. Breast Cancer Res. 2016;18:49.ArticlePubMedPubMed CentralCASGoogle Scholar
Rygiel TP, Stolte EH, de Ruiter T, van de Weijer ML, Meyaard L. Tumor-expressed collagens can modulate immune cell function through the inhibitory collagen receptor LAIR-1. Mol Immunol. 2011;49:402–6.ArticleCASPubMedGoogle Scholar
Sprague L, Muccioli M, Pate M, Singh M, Xiong C, Ostermann A, Niese B, Li Y, Li Y, Courreges MC, Benencia F. Dendritic cells: in vitro culture in two- and three-dimensional collagen systems and expression of collagen receptors in tumors and atherosclerotic microenvironments. Exp Cell Res. 2014;323:7–27.ArticleCASPubMedGoogle Scholar
Okumura T, Ohuchida K, Kibe S, Iwamoto C, Ando Y, Takesue S, Nakayama H, Abe T, Endo S, Koikawa K, et al. Adipose tissue-derived stromal cells are sources of cancer-associated fibroblasts and enhance tumor progression by dense collagen matrix. Int J Cancer. 2019;144:1401–13.ArticleCASPubMedGoogle Scholar
Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16:582–98.ArticleCASPubMedGoogle Scholar
Jones CE, Hammer AM, Cho Y, Sizemore GM, Cukierman E, Yee LD, Ghadiali SN, Ostrowski MC, Leight JL. Stromal PTEN regulates extracellular matrix organization in the mammary gland. Neoplasia. 2019;21:132–45.ArticleCASPubMedGoogle Scholar
Miyashita T, Omori T, Nakamura H, Sugano M, Neri S, Fujii S, Hashimoto H, Tsuboi M, Ochiai A, Ishii G. Spatiotemporal characteristics of fibroblasts-dependent cancer cell invasion. J Cancer Res Clin Oncol. 2019;145:373–81.ArticleCASPubMedGoogle Scholar
Keating M, Kurup A, Alvarez-Elizondo M, Levine AJ, Botvinick E. Spatial distributions of pericellular stiffness in natural extracellular matrices are dependent on cell-mediated proteolysis and contractility. Acta Biomater. 2017;57:304–12.ArticleCASPubMedPubMed CentralGoogle Scholar
Yang YL, Sun C, Wilhelm ME, Fox LJ, Zhu J, Kaufman LJ. Influence of chondroitin sulfate and hyaluronic acid on structure, mechanical properties, and glioma invasion of collagen I gels. Biomaterials. 2011;32:7932–40.ArticleCASPubMedPubMed CentralGoogle Scholar
Miroshnikova YA, Rozenberg GI, Cassereau L, Pickup M, Mouw JK, Ou G, Templeman KL, Hannachi EI, Gooch KJ, Sarang-Sieminski AL, et al. alpha5beta1-Integrin promotes tension-dependent mammary epithelial cell invasion by engaging the fibronectin synergy site. Mol Biol Cell. 2017;28:2958–77.ArticleCASPubMedPubMed CentralGoogle Scholar
Biondani G, Zeeberg K, Greco MR, Cannone S, Dando I, Dalla Pozza E, Mastrodonato M, Forciniti S, Casavola V, Palmieri M, et al. Extracellular matrix composition modulates PDAC parenchymal and stem cell plasticity and behavior through the secretome. FEBS J. 2018;285:2104–24.ArticleCASPubMedGoogle Scholar
Vellinga TT, den Uil S, Rinkes IH, Marvin D, Ponsioen B, Alvarez-Varela A, Fatrai S, Scheele C, Zwijnenburg DA, Snippert H, et al. Collagen-rich stroma in aggressive colon tumors induces mesenchymal gene expression and tumor cell invasion. Oncogene. 2016;35:5263–71.ArticleCASPubMedGoogle Scholar
Kubow KE, Vukmirovic R, Zhe L, Klotzsch E, Smith ML, Gourdon D, Luna S, Vogel V. Mechanical forces regulate the interactions of fibronectin and collagen I in extracellular matrix. Nat Commun. 2015;6:8026.ArticleCASPubMedGoogle Scholar
Wang K, Wu F, Seo BR, Fischbach C, Chen W, Hsu L, Gourdon D. Breast cancer cells alter the dynamics of stromal fibronectin-collagen interactions. Matrix Biol. 2017;60–61:86–95.ArticlePubMedCASGoogle Scholar
Chang KC, Lu YC, Lin MJ, Chen HY, Jin YT. Desmoplastic tumour-associated stroma versus neural tissue in central nervous system metastasis: effects of different microenvironments on tumour growth. Histopathology. 2011;59:31–9.ArticlePubMedGoogle Scholar
Hurskainen M, Ruggiero F, Hagg P, Pihlajaniemi T, Huhtala P. Recombinant human collagen XV regulates cell adhesion and migration. J Biol Chem. 2010;285:5258–65.ArticleCASPubMedGoogle Scholar
Jang M, Koh I, Lee JE, Lim JY, Cheong JH, Kim P. Increased extracellular matrix density disrupts E-cadherin/beta-catenin complex in gastric cancer cells. Biomater Sci. 2018;6:2704–13.ArticleCASPubMedGoogle Scholar
Wegner CS, Gaustad JV, Andersen LM, Simonsen TG, Rofstad EK. Diffusion-weighted and dynamic contrast-enhanced MRI of pancreatic adenocarcinoma xenografts: associations with tumor differentiation and collagen content. J Transl Med. 2016;14:161.ArticlePubMedPubMed CentralCASGoogle Scholar
Mammoto T, Jiang A, Jiang E, Panigrahy D, Kieran MW, Mammoto A. Role of collagen matrix in tumor angiogenesis and glioblastoma multiforme progression. Am J Pathol. 2013;183:1293–305.ArticleCASPubMedPubMed CentralGoogle Scholar
Thangavelu PU, Krenacs T, Dray E, Duijf PH. In epithelial cancers, aberrant COL17A1 promoter methylation predicts its misexpression and increased invasion. Clin Epigenetics. 2016;8:120.ArticlePubMedPubMed CentralCASGoogle Scholar
Provenzano PP, Eliceiri KW, Campbell JM, Inman DR, White JG, Keely PJ. Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 2006;4:38.ArticlePubMedPubMed CentralCASGoogle Scholar
Conklin MW, Gangnon RE, Sprague BL, Van Gemert L, Hampton JM, Eliceiri KW, Bredfeldt JS, Liu Y, Surachaicharn N, Newcomb PA, et al. Collagen alignment as a predictor of recurrence after ductal carcinoma in situ. Cancer Epidemiol Biomarkers Prev. 2018;27:138–45.ArticleCASPubMedGoogle Scholar
Conklin MW, Eickhoff JC, Riching KM, Pehlke CA, Eliceiri KW, Provenzano PP, Friedl A, Keely PJ. Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am J Pathol. 2011;178:1221–32.ArticlePubMedPubMed CentralGoogle Scholar
Humphries BA, Buschhaus JM, Chen YC, Haley HR, Qyli T, Chiang B, Shen N, Rajendran S, Cutter A, Cheng YH, et al. Plasminogen activator inhibitor 1 (PAI1) promotes actin cytoskeleton reorganization and glycolytic metabolism in triple-negative breast cancer. Mol Cancer Res. 2019;17:1142–54.PubMedPubMed CentralGoogle Scholar
Provenzano PP, Inman DR, Eliceiri KW, Keely PJ. Matrix density-induced mechanoregulation of breast cell phenotype, signaling and gene expression through a FAK-ERK linkage. Oncogene. 2009;28:4326–43.ArticleCASPubMedPubMed CentralGoogle Scholar
Samuel MS, Lopez JI, McGhee EJ, Croft DR, Strachan D, Timpson P, Munro J, Schroder E, Zhou J, Brunton VG, et al. Actomyosin-mediated cellular tension drives increased tissue stiffness and beta-catenin activation to induce epidermal hyperplasia and tumor growth. Cancer Cell. 2011;19:776–91.ArticleCASPubMedPubMed CentralGoogle Scholar
Ohlund D, Franklin O, Lundberg E, Lundin C, Sund M. Type IV collagen stimulates pancreatic cancer cell proliferation, migration, and inhibits apoptosis through an autocrine loop. BMC Cancer. 2013;13:154.ArticlePubMedPubMed CentralCASGoogle Scholar
Berchtold S, Grunwald B, Kruger A, Reithmeier A, Hahl T, Cheng T, Feuchtinger A, Born D, Erkan M, Kleeff J, Esposito I. Collagen type V promotes the malignant phenotype of pancreatic ductal adenocarcinoma. Cancer Lett. 2015;356:721–32.ArticleCASPubMedGoogle Scholar
Ueno H, Konishi T, Ishikawa Y, Shimazaki H, Ueno M, Aosasa S, Saiura A, Hase K, Yamamoto J. Histologic categorization of fibrotic cancer stroma in the primary tumor is an independent prognostic index in resectable colorectal liver metastasis. Am J Surg Pathol. 2014;38:1380–6.ArticlePubMedGoogle Scholar
Dayal JH, Cole CL, Pourreyron C, Watt SA, Lim YZ, Salas-Alanis JC, Murrell DF, McGrath JA, Stieger B, Jahoda C, et al. Type VII collagen regulates expression of OATP1B3, promotes front-to-rear polarity and increases structural organisation in 3D spheroid cultures of RDEB tumour keratinocytes. J Cell Sci. 2014;127:740–51.ArticleCASPubMedPubMed CentralGoogle Scholar
Zhou ZH, Ji CD, Xiao HL, Zhao HB, Cui YH, Bian XW. Reorganized collagen in the tumor microenvironment of gastric cancer and its association with prognosis. J Cancer. 2017;8:1466–76.ArticlePubMedPubMed CentralCASGoogle Scholar
Sapudom J, Kalbitzer L, Wu X, Martin S, Kroy K, Pompe T. Fibril bending stiffness of 3D collagen matrices instructs spreading and clustering of invasive and non-invasive breast cancer cells. Biomaterials. 2019;193:47–57.ArticleCASPubMedGoogle Scholar
Brauchle E, Kasper J, Daum R, Schierbaum N, Falch C, Kirschniak A, Schaffer TE, Schenke-Layland K. Biomechanical and biomolecular characterization of extracellular matrix structures in human colon carcinomas. Matrix Biol. 2018;68–69:180–93.ArticlePubMedCASGoogle Scholar
Phadnis SV, Atilade A, Bowring J, Kyrgiou M, Young MP, Evans H, Paraskevaidis E, Walker P. Regeneration of cervix after excisional treatment for cervical intraepithelial neoplasia: a study of collagen distribution. BJOG. 2011;118:1585–91.ArticleCASPubMedGoogle Scholar
Senthebane DA, Jonker T, Rowe A, Thomford NE, Munro D, Dandara C, Wonkam A, Govender D, Calder B, Soares NC, et al. The role of tumor microenvironment in chemoresistance: 3D extracellular matrices as accomplices. Int J Mol Sci. 2018;19:E2861.ArticlePubMedCASGoogle Scholar
Liu L, Zhang SX, Liao W, Farhoodi HP, Wong CW, Chen CC, Segaliny AI, Chacko JV, Nguyen LP, Lu M, et al. Mechanoresponsive stem cells to target cancer metastases through biophysical cues. Sci Transl Med. 2017;9:eaan2966.ArticlePubMedPubMed CentralCASGoogle Scholar
Li X, Shepard HM, Cowell JA, Zhao C, Osgood RJ, Rosengren S, Blouw B, Garrovillo SA, Pagel MD, Whatcott CJ, et al. Parallel accumulation of tumor hyaluronan, collagen, and other drivers of tumor progression. Clin Cancer Res. 2018;24:4798–807.ArticleCASPubMedPubMed CentralGoogle Scholar
Jakubzig B, Baltes F, Henze S, Schlesinger M, Bendas G. Mechanisms of matrix-induced chemoresistance of breast cancer cells-deciphering novel potential targets for a cell sensitization. Cancers (Basel). 2018;10:E495.ArticleGoogle Scholar
Hu B, Mao Z, Jiang X, He D, Wang Z, Wang X, Zhu Y, Wang H. Role of TGF-beta1/Smad3-mediated fibrosis in drug resistance mechanism of prolactinoma. Brain Res. 2018;1698:204–12.ArticleCASPubMedGoogle Scholar
Sterzynska K, Klejewski A, Wojtowicz K, Swierczewska M, Nowacka M, Kazmierczak D, Andrzejewska M, Rusek D, Brazert M, Brazert J, et al. Mutual expression of ALDH1A1, LOX, and collagens in ovarian cancer cell lines as combined CSCs- and ECM-related models of drug resistance development. Int J Mol Sci. 2018;20:E54.ArticlePubMedCASGoogle Scholar
Gurler H, Yu Y, Choi J, Kajdacsy-Balla AA, Barbolina MV. Three-dimensional collagen type I matrix up-regulates nuclear isoforms of the microtubule associated protein tau implicated in resistance to paclitaxel therapy in ovarian carcinoma. Int J Mol Sci. 2015;16:3419–33.ArticleCASPubMedPubMed CentralGoogle Scholar
Krasny L, Shimony N, Tzukert K, Gorodetsky R, Lecht S, Nettelbeck DM, Haviv YS. An in vitro tumour microenvironment model using adhesion to type I collagen reveals Akt-dependent radiation resistance in renal cancer cells. Nephrol Dial Transplant. 2010;25:373–80.ArticleCASPubMedGoogle Scholar
Januchowski R, Swierczewska M, Sterzynska K, Wojtowicz K, Nowicki M, Zabel M. Increased expression of several collagen genes is associated with drug resistance in ovarian cancer cell lines. J Cancer. 2016;7:1295–310.ArticleCASPubMedPubMed CentralGoogle Scholar
Goncalves-Ribeiro S, Sanz-Pamplona R, Vidal A, Sanjuan X, Guillen Diaz-Maroto N, Soriano A, Guardiola J, Albert N, Martinez-Villacampa M, Lopez I, et al. Prediction of pathological response to neoadjuvant treatment in rectal cancer with a two-protein immunohistochemical score derived from stromal gene-profiling. Ann Oncol. 2017;28:2160–8.ArticleCASPubMedGoogle Scholar
Burnier JV, Wang N, Michel RP, Hassanain M, Li S, Lu Y, Metrakos P, Antecka E, Burnier MN, Ponton A, et al. Type IV collagen-initiated signals provide survival and growth cues required for liver metastasis. Oncogene. 2011;30:3766–83.ArticleCASPubMedGoogle Scholar
Angenendt L, Mikesch JH, Gorlich D, Busch A, Arnhold I, Rudack C, Hartmann W, Wardelmann E, Berdel WE, Stenner M, et al. Stromal collagen type VI associates with features of malignancy and predicts poor prognosis in salivary gland cancer. Cell Oncol (Dordr). 2018;41:517–25.ArticleCASGoogle Scholar
Rolff HC, Christensen IJ, Vainer B, Svendsen LB, Eefsen RL, Wilhelmsen M, Lund IK, Hoyer-Hansen G, Nielsen HJ, Illemann M, Danish Collaborative Research Group on Colorectal C. The prognostic and predictive value of soluble type IV collagen in colorectal cancer: a retrospective multicenter study. Clin Cancer Res. 2016;22:2427–34.ArticleCASPubMedGoogle Scholar
Aguilera KY, Rivera LB, Hur H, Carbon JG, Toombs JE, Goldstein CD, Dellinger MT, Castrillon DH, Brekken RA. Collagen signaling enhances tumor progression after anti-VEGF therapy in a murine model of pancreatic ductal adenocarcinoma. Cancer Res. 2014;74:1032–44.ArticleCASPubMedGoogle Scholar
Thaci B, Ulasov IV, Ahmed AU, Ferguson SD, Han Y, Lesniak MS. Anti-angiogenic therapy increases intratumoral adenovirus distribution by inducing collagen degradation. Gene Ther. 2013;20:318–27.ArticleCASPubMedGoogle Scholar
Rahbari NN, Kedrin D, Incio J, Liu H, Ho WW, Nia HT, Edrich CM, Jung K, Daubriac J, Chen I, et al. Anti-VEGF therapy induces ECM remodeling and mechanical barriers to therapy in colorectal cancer liver metastases. Sci Transl Med. 2016;8:360ra135.ArticlePubMedPubMed CentralCASGoogle Scholar
Wu YH, Huang YF, Chang TH, Chou CY. Activation of TWIST1 by COL11A1 promotes chemoresistance and inhibits apoptosis in ovarian cancer cells by modulating NF-kappaB-mediated IKKbeta expression. Int J Cancer. 2017;141:2305–17.ArticleCASPubMedGoogle Scholar
Wang H, Mislati R, Ahmed R, Vincent P, Nwabunwanne SF, Gunn JR, Pogue BW, Doyley MM. Elastography can map the local inverse relationship between shear modulus and drug delivery within the pancreatic ductal adenocarcinoma microenvironment. Clin Cancer Res. 2019;25:2136–43.ArticlePubMedGoogle Scholar
Bennink LL, Li Y, Kim B, Shin IJ, San BH, Zangari M, Yoon D, Yu SM. Visualizing collagen proteolysis by peptide hybridization: from 3D cell culture to in vivo imaging. Biomaterials. 2018;183:67–76.ArticleCASPubMedGoogle Scholar
Golaraei A, Mirsanaye K, Ro Y, Krouglov S, Akens MK, Wilson BC, Barzda V. Collagen chirality and three-dimensional orientation studied with polarimetric second-harmonic generation microscopy. J Biophotonics. 2019;12:e201800241.ArticlePubMedCASGoogle Scholar
Montesi SB, Desogere P, Fuchs BC, Caravan P. Molecular imaging of fibrosis: recent advances and future directions. J Clin Invest. 2019;129:24–33.ArticlePubMedPubMed CentralGoogle Scholar
Aoyagi T, Shuto K, Okazumi S, Hayano K, Satoh A, Saitoh H, Shimada H, Nabeya Y, Kazama T, Matsubara H. Apparent diffusion coefficient correlation with oesophageal tumour stroma and angiogenesis. Eur Radiol. 2012;22:1172–7.ArticlePubMedGoogle Scholar
Paidi SK, Diaz PM, Dadgar S, Jenkins SV, Quick CM, Griffin RJ, Dings RPM, Rajaram N, Barman I. Label-free Raman spectroscopy reveals signatures of radiation resistance in the tumor microenvironment. Cancer Res. 2019;79:2054–64.ArticleCASPubMedPubMed CentralGoogle Scholar
Chronopoulos A, Robinson B, Sarper M, Cortes E, Auernheimer V, Lachowski D, Attwood S, Garcia R, Ghassemi S, Fabry B, Del Rio Hernandez A. ATRA mechanically reprograms pancreatic stellate cells to suppress matrix remodelling and inhibit cancer cell invasion. Nat Commun. 2016;7:12630.ArticlePubMedPubMed CentralGoogle Scholar
Bleul T, Ruhl R, Bulashevska S, Karakhanova S, Werner J, Bazhin AV. Reduced retinoids and retinoid receptors’ expression in pancreatic cancer: a link to patient survival. Mol Carcinog. 2015;54:870–9.ArticleCASPubMedGoogle Scholar
Bynigeri RR, Jakkampudi A, Jangala R, Subramanyam C, Sasikala M, Rao GV, Reddy DN, Talukdar R. Pancreatic stellate cell: Pandora’s box for pancreatic disease biology. World J Gastroenterol. 2017;23:382–405.ArticleCASPubMedPubMed CentralGoogle Scholar
Hummel D, Aggarwal A, Borka K, Bajna E, Kallay E, Horvath HC. The vitamin D system is deregulated in pancreatic diseases. J Steroid Biochem Mol Biol. 2014;144(Pt B):402–9.ArticleCASPubMedPubMed CentralGoogle Scholar
Sherman MH, Yu RT, Engle DD, Ding N, Atkins AR, Tiriac H, Collisson EA, Connor F, Van Dyke T, Kozlov S, et al. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell. 2014;159:80–93.ArticleCASPubMedPubMed CentralGoogle Scholar
Karasic TB, O’Hara MH, Loaiza-Bonilla A, Reiss KA, Teitelbaum UR, Borazanci E, De Jesus-Acosta A, Redlinger C, Burrell JA, Laheru DA, et al. Effect of Gemcitabine and nab-paclitaxel with or without hydroxychloroquine on patients with advanced pancreatic cancer: a phase 2 randomized clinical trial. JAMA Oncol. 2019. https://doi.org/10.1001/jamaoncol.2019.0684.ArticlePubMedPubMed CentralGoogle Scholar
Awasthi N, Mikels-Vigdal AJ, Stefanutti E, Schwarz MA, Monahan S, Smith V, Schwarz RE. Therapeutic efficacy of anti-MMP9 antibody in combination with nab-paclitaxel-based chemotherapy in pre-clinical models of pancreatic cancer. J Cell Mol Med. 2019;23:3878–87.ArticleCASPubMedPubMed CentralGoogle Scholar
Catenacci DV, Junttila MR, Karrison T, Bahary N, Horiba MN, Nattam SR, Marsh R, Wallace J, Kozloff M, Rajdev L, et al. Randomized phase Ib/II study of gemcitabine plus placebo or vismodegib, a hedgehog pathway inhibitor, in patients with metastatic pancreatic cancer. J Clin Oncol. 2015;33:4284–92.ArticleCASPubMedPubMed CentralGoogle Scholar
Elahi-Gedwillo KY, Carlson M, Zettervall J, Provenzano PP. Antifibrotic therapy disrupts stromal barriers and modulates the immune landscape in pancreatic ductal adenocarcinoma. Cancer Res. 2019;79:372–86.ArticleCASPubMedGoogle Scholar
Seniutkin O, Furuya S, Luo YS, Cichocki JA, Fukushima H, Kato Y, Sugimoto H, Matsumoto T, Uehara T, Rusyn I. Effects of pirfenidone in acute and sub-chronic liver fibrosis, and an initiation-promotion cancer model in the mouse. Toxicol Appl Pharmacol. 2018;339:1–9.ArticleCASPubMedGoogle Scholar
Pan A, Wang Z, Chen B, Dai W, Zhang H, He B, Wang X, Wang Y, Zhang Q. Localized co-delivery of collagenase and trastuzumab by thermosensitive hydrogels for enhanced antitumor efficacy in human breast xenograft. Drug Deliv. 2018;25:1495–503.ArticleCASPubMedPubMed CentralGoogle Scholar
Dong X, Liu HJ, Feng HY, Yang SC, Liu XL, Lai X, Lu Q, Lovell JF, Chen HZ, Fang C. Enhanced drug delivery by nanoscale integration of a nitric oxide donor to induce tumor collagen depletion. Nano Lett. 2019;19:997–1008.ArticlePubMedCASGoogle Scholar
Li J, Huang J, Ao Y, Li S, Miao Y, Yu Z, Zhu L, Lan X, Zhu Y, Zhang Y, Yang X. Synergizing upconversion nanophotosensitizers with hyperbaric oxygen to remodel the extracellular matrix for enhanced photodynamic cancer therapy. ACS Appl Mater Interfaces. 2018;10:22985–96.ArticleCASPubMedGoogle Scholar
Happold C, Gorlia T, Chinot O, Gilbert MR, Nabors LB, Wick W, Pugh SL, Hegi M, Cloughesy T, Roth P, et al. Does valproic acid or levetiracetam improve survival in glioblastoma? A pooled analysis of prospective clinical trials in newly diagnosed glioblastoma. J Clin Oncol. 2016;34:731–9.ArticleCASPubMedPubMed CentralGoogle Scholar
Wang L, Liu X, Zhou Q, Sui M, Lu Z, Zhou Z, Tang J, Miao Y, Zheng M, Wang W, Shen Y. Terminating the criminal collaboration in pancreatic cancer: nanoparticle-based synergistic therapy for overcoming fibroblast-induced drug resistance. Biomaterials. 2017;144:105–18.ArticleCASPubMedGoogle Scholar
Cun X, Ruan S, Chen J, Zhang L, Li J, He Q, Gao H. A dual strategy to improve the penetration and treatment of breast cancer by combining shrinking nanoparticles with collagen depletion by losartan. Acta Biomater. 2016;31:186–96.ArticleCASPubMedGoogle Scholar
Tang Y, Liu Y, Wang S, Tian Y, Li Y, Teng Z, Lu G. Depletion of collagen by losartan to improve tumor accumulation and therapeutic efficacy of photodynamic nanoplatforms. Drug Deliv Transl Res. 2019;9:615–24.ArticleCASPubMedGoogle Scholar
Shen H, Gao Q, Ye Q, Yang S, Wu Y, Huang Q, Wang X, Sun Z. Peritumoral implantation of hydrogel-containing nanoparticles and losartan for enhanced nanoparticle penetration and antitumor effect. Int J Nanomed. 2018;13:7409–26.ArticleCASGoogle Scholar
Whatcott CJ, Han H, Von Hoff DD. Orchestrating the tumor microenvironment to improve survival for patients with pancreatic cancer: normalization, not destruction. Cancer J. 2015;21:299–306.ArticleCASPubMedPubMed CentralGoogle Scholar
Li J, Xie C, Huang J, Jiang Y, Miao Q, Pu K. Semiconducting polymer nanoenzymes with photothermic activity for enhanced cancer therapy. Angew Chem Int Ed Engl. 2018;57:3995–8.ArticleCASPubMedGoogle Scholar
Kolosnjaj-Tabi J, Marangon I, Nicolas-Boluda A, Silva AKA, Gazeau F. Nanoparticle-based hyperthermia, a local treatment modulating the tumor extracellular matrix. Pharmacol Res. 2017;126:123–37.ArticleCASPubMedGoogle Scholar
Ayalew L, Acuna J, Urfano SF, Morfin C, Sablan A, Oh M, Gamboa A, Slowinska K. Conjugation of paclitaxel to hybrid peptide carrier and biological evaluation in jurkat and A549 cancer cell lines. ACS Med Chem Lett. 2017;8:814–9.ArticleCASPubMedPubMed CentralGoogle Scholar
Liang H, Li X, Chen B, Wang B, Zhao Y, Zhuang Y, Shen H, Zhang Z, Dai J. A collagen-binding EGFR single-chain Fv antibody fragment for the targeted cancer therapy. J Control Release. 2015;209:101–9.ArticleCASPubMedGoogle Scholar
Ishihara J, Ishihara A, Sasaki K, Lee SS, Williford JM, Yasui M, Abe H, Potin L, Hosseinchi P, Fukunaga K, et al. Targeted antibody and cytokine cancer immunotherapies through collagen affinity. Sci Transl Med. 2019;11:eaau3259.ArticleCASPubMedPubMed CentralGoogle Scholar
Yasunaga M, Manabe S, Tarin D, Matsumura Y. Cancer-stroma targeting therapy by cytotoxic immunoconjugate bound to the collagen 4 network in the tumor tissue. Bioconjug Chem. 2011;22:1776–83.ArticleCASPubMedGoogle Scholar
Blackburn JS, Liu I, Coon CI, Brinckerhoff CE. A matrix metalloproteinase-1/protease activated receptor-1 signaling axis promotes melanoma invasion and metastasis. Oncogene. 2009;28:4237–48.ArticleCASPubMedPubMed CentralGoogle Scholar
Devy J, Duca L, Cantarelli B, Joseph-Pietras D, Scandolera A, Rusciani A, Parent L, Thevenard J, Pasco SB, Tarpin M, et al. Elastin-derived peptides enhance melanoma growth in vivo by upregulating the activation of Mcol-A (MMP-1) collagenase. Br J Cancer. 2010;103:1562–70.ArticleCASPubMedPubMed CentralGoogle Scholar
Rider L, Oladimeji P, Diakonova M. PAK1 regulates breast cancer cell invasion through secretion of matrix metalloproteinases in response to prolactin and three-dimensional collagen IV. Mol Endocrinol. 2013;27:1048–64.ArticleCASPubMedPubMed CentralGoogle Scholar
Elenjord R, Allen JB, Johansen HT, Kildalsen H, Svineng G, Maelandsmo GM, Loennechen T, Winberg JO. Collagen I regulates matrix metalloproteinase-2 activation in osteosarcoma cells independent of S100A4. FEBS J. 2009;276:5275–86.ArticleCASPubMedGoogle Scholar
Freise C, Erben U, Muche M, Farndale R, Zeitz M, Somasundaram R, Ruehl M. The alpha 2 chain of collagen type VI sequesters latent proforms of matrix-metalloproteinases and modulates their activation and activity. Matrix Biol. 2009;28:480–9.ArticleCASPubMedGoogle Scholar
Bates AL, Pickup MW, Hallett MA, Dozier EA, Thomas S, Fingleton B. Stromal matrix metalloproteinase 2 regulates collagen expression and promotes the outgrowth of experimental metastases. J Pathol. 2015;235:773–83.ArticleCASPubMedPubMed CentralGoogle Scholar
Koikawa K, Ohuchida K, Ando Y, Kibe S, Nakayama H, Takesue S, Endo S, Abe T, Okumura T, Iwamoto C, et al. Basement membrane destruction by pancreatic stellate cells leads to local invasion in pancreatic ductal adenocarcinoma. Cancer Lett. 2018;425:65–77.ArticleCASPubMedGoogle Scholar
Sandri S, Faiao-Flores F, Tiago M, Pennacchi PC, Massaro RR, Alves-Fernandes DK, Berardinelli GN, Evangelista AF, de Lima Vazquez V, Reis RM, Maria-Engler SS. Vemurafenib resistance increases melanoma invasiveness and modulates the tumor microenvironment by MMP-2 upregulation. Pharmacol Res. 2016;111:523–33.ArticleCASPubMedGoogle Scholar
Mahabir R, Tanino M, Elmansuri A, Wang L, Kimura T, Itoh T, Ohba Y, Nishihara H, Shirato H, Tsuda M, Tanaka S. Sustained elevation of Snail promotes glial-mesenchymal transition after irradiation in malignant glioma. Neuro Oncol. 2014;16:671–85.ArticleCASPubMedGoogle Scholar
Correia AL, Mori H, Chen EI, Schmitt FC, Bissell MJ. The hemopexin domain of MMP3 is responsible for mammary epithelial invasion and morphogenesis through extracellular interaction with HSP90beta. Genes Dev. 2013;27:805–17.ArticleCASPubMedPubMed CentralGoogle Scholar
Prakobwong S, Charoensuk L, Hiraku Y, Pinlaor P, Pairojkul C, Mairiang E, Sithithaworn P, Yongvanit P, Khuntikeo N, Pinlaor S. Plasma hydroxyproline, MMP-7 and collagen I as novel predictive risk markers of hepatobiliary disease-associated cholangiocarcinoma. Int J Cancer. 2012;131:E416–24.ArticleCASPubMedGoogle Scholar
Orgaz JL, Pandya P, Dalmeida R, Karagiannis P, Sanchez-Laorden B, Viros A, Albrengues J, Nestle FO, Ridley AJ, Gaggioli C, et al. Diverse matrix metalloproteinase functions regulate cancer amoeboid migration. Nat Commun. 2014;5:4255.ArticleCASPubMedGoogle Scholar
Prakobwong S, Yongvanit P, Hiraku Y, Pairojkul C, Sithithaworn P, Pinlaor P, Pinlaor S. Involvement of MMP-9 in peribiliary fibrosis and cholangiocarcinogenesis via Rac1-dependent DNA damage in a hamster model. Int J Cancer. 2010;127:2576–87.ArticleCASPubMedGoogle Scholar
Cho K, Matsuda Y, Ueda J, Uchida E, Naito Z, Ishiwata T. Keratinocyte growth factor induces matrix metalloproteinase-9 expression and correlates with venous invasion in pancreatic cancer. Int J Oncol. 2012;40:1040–8.ArticleCASPubMedGoogle Scholar
Wilkins-Port CE, Ye Q, Mazurkiewicz JE, Higgins PJ. TGF-beta1 + EGF-initiated invasive potential in transformed human keratinocytes is coupled to a plasmin/MMP-10/MMP-1-dependent collagen remodeling axis: role for PAI-1. Cancer Res. 2009;69:4081–91.ArticleCASPubMedPubMed CentralGoogle Scholar
Krantz SB, Shields MA, Dangi-Garimella S, Cheon EC, Barron MR, Hwang RF, Rao MS, Grippo PJ, Bentrem DJ, Munshi HG. MT1-MMP cooperates with Kras(G12D) to promote pancreatic fibrosis through increased TGF-beta signaling. Mol Cancer Res. 2011;9:1294–304.ArticleCASPubMedPubMed CentralGoogle Scholar
Borrirukwanit K, Pavasant P, Blick T, Lafleur MA, Thompson EW. High threshold of beta1 integrin inhibition required to block collagen I-induced membrane type-1 matrix metalloproteinase (MT1-MMP) activation of matrix metalloproteinase 2 (MMP-2). Cancer Cell Int. 2014;14:99.ArticlePubMedPubMed CentralCASGoogle Scholar
Maquoi E, Assent D, Detilleux J, Pequeux C, Foidart JM, Noel A. MT1-MMP protects breast carcinoma cells against type I collagen-induced apoptosis. Oncogene. 2012;31:480–93.ArticleCASPubMedGoogle Scholar
Moss NM, Liu Y, Johnson JJ, Debiase P, Jones J, Hudson LG, Munshi HG, Stack MS. Epidermal growth factor receptor-mediated membrane type 1 matrix metalloproteinase endocytosis regulates the transition between invasive versus expansive growth of ovarian carcinoma cells in three-dimensional collagen. Mol Cancer Res. 2009;7:809–20.ArticleCASPubMedPubMed CentralGoogle Scholar
Tatti O, Gucciardo E, Pekkonen P, Holopainen T, Louhimo R, Repo P, Maliniemi P, Lohi J, Rantanen V, Hautaniemi S, et al. MMP16 mediates a proteolytic switch to promote cell-cell adhesion, collagen alignment, and lymphatic invasion in melanoma. Cancer Res. 2015;75:2083–94.ArticleCASPubMedGoogle Scholar
Rodgers UR, Kevorkian L, Surridge AK, Waters JG, Swingler TE, Culley K, Illman S, Lohi J, Parker AE, Clark IM. Expression and function of matrix metalloproteinase (MMP)-28. Matrix Biol. 2009;28:263–72.ArticleCASPubMedPubMed CentralGoogle Scholar
Zou X, Feng B, Dong T, Yan G, Tan B, Shen H, Huang A, Zhang X, Zhang M, Yang P, et al. Up-regulation of type I collagen during tumorigenesis of colorectal cancer revealed by quantitative proteomic analysis. J Proteomics. 2013;94:473–85.ArticleCASPubMedGoogle Scholar
Penet MF, Kakkad S, Pathak AP, Krishnamachary B, Mironchik Y, Raman V, Solaiyappan M, Bhujwalla ZM. Structure and function of a prostate cancer dissemination-permissive extracellular matrix. Clin Cancer Res. 2017;23:2245–54.ArticleCASPubMedGoogle Scholar
Xiong G, Deng L, Zhu J, Rychahou PG, Xu R. Prolyl-4-hydroxylase alpha subunit 2 promotes breast cancer progression and metastasis by regulating collagen deposition. BMC Cancer. 2014;14:1.ArticlePubMedPubMed CentralCASGoogle Scholar
Yu Y, Liu D, Liu Z, Li S, Ge Y, Sun W, Liu B. The inhibitory effects of COL1A2 on colorectal cancer cell proliferation, migration, and invasion. J Cancer. 2018;9:2953–62.ArticlePubMedPubMed CentralCASGoogle Scholar
Ji J, Zhao L, Budhu A, Forgues M, Jia HL, Qin LX, Ye QH, Yu J, Shi X, Tang ZY, Wang XW. Let-7g targets collagen type I alpha2 and inhibits cell migration in hepatocellular carcinoma. J Hepatol. 2010;52:690–7.ArticleCASPubMedPubMed CentralGoogle Scholar
Tamiya M, Kobayashi M, Morimura O, Yasue T, Nakasuji T, Satomu M, Kohei O, Takayuki S, Morishita N, Suzuki H, et al. Clinical significance of the serum crosslinked N-telopeptide of type I collagen as a prognostic marker for non-small-cell lung cancer. Clin Lung Cancer. 2013;14:50–4.ArticleCASPubMedGoogle Scholar
Nurmenniemi S, Koivula MK, Nyberg P, Tervahartiala T, Sorsa T, Mattila PS, Salo T, Risteli J. Type I and III collagen degradation products in serum predict patient survival in head and neck squamous cell carcinoma. Oral Oncol. 2012;48:136–40.ArticleCASPubMedGoogle Scholar
Imamura M, Nishimukai A, Higuchi T, Ozawa H, Yanai A, Miyagawa Y, Murase K, Sakita I, Hatada T, Takatsuka Y, et al. High levels at baseline of serum pyridinoline crosslinked carboxyterminal telopeptide of type I collagen are associated with worse prognosis for breast cancer patients. Breast Cancer Res Treat. 2015;154:521–31.ArticleCASPubMedGoogle Scholar
Ferreira AR, Alho I, Shan N, Matias M, Faria M, Casimiro S, Leitzel K, Ali S, Lipton A, Costa L. N-telopeptide of type i collagen long-term dynamics in breast cancer patients with bone metastases: clinical outcomes and influence of extraskeletal metastases. Oncologist. 2016;21:1418–26.ArticleCASPubMedPubMed CentralGoogle Scholar
Ganapathi MK, Jones WD, Sehouli J, Michener CM, Braicu IE, Norris EJ, Biscotti CV, Vaziri SA, Ganapathi RN. Expression profile of COL2A1 and the pseudogene SLC6A10P predicts tumor recurrence in high-grade serous ovarian cancer. Int J Cancer. 2016;138:679–88.ArticleCASPubMedGoogle Scholar
Tarpey PS, Behjati S, Cooke SL, Van Loo P, Wedge DC, Pillay N, Marshall J, O’Meara S, Davies H, Nik-Zainal S, et al. Frequent mutation of the major cartilage collagen gene COL2A1 in chondrosarcoma. Nat Genet. 2013;45:923–6.ArticleCASPubMedPubMed CentralGoogle Scholar
Wang Y, Resnick MB, Lu S, Hui Y, Brodsky AS, Yang D, Yakirevich E, Wang L. Collagen type III alpha1 as a useful diagnostic immunohistochemical marker for fibroepithelial lesions of the breast. Hum Pathol. 2016;57:176–81.ArticleCASPubMedGoogle Scholar
Ando H, Aihara R, Ohno T, Ogata K, Mochiki E, Kuwano H. Prognostic significance of the expression of MUC1 and collagen type IV in advanced gastric carcinoma. Br J Surg. 2009;96:901–9.ArticleCASPubMedGoogle Scholar
Fan HX, Li HX, Chen D, Gao ZX, Zheng JH. Changes in the expression of MMP2, MMP9, and ColIV in stromal cells in oral squamous tongue cell carcinoma: relationships and prognostic implications. J Exp Clin Cancer Res. 2012;31:90.ArticleCASPubMedPubMed CentralGoogle Scholar
Nystrom H, Naredi P, Hafstrom L, Sund M. Type IV collagen as a tumour marker for colorectal liver metastases. Eur J Surg Oncol. 2011;37:611–7.ArticleCASPubMedGoogle Scholar
Shimizu A, Kobayashi A, Yokoyama T, Motoyama H, Sakai H, Kitagawa N, Notake T, Shirota T, Fukushima K, Miyagawa SI. Correlation between the serum levels of type IV collagen 7 s domain and the risk of intractable ascites following liver resection for hepatocellular carcinoma: a propensity score-matched analysis. Surgery. 2016;160:1244–55.ArticlePubMedGoogle Scholar
Pan Z, Li L, Fang Q, Zhang Y, Hu X, Qian Y, Huang P. Analysis of dynamic molecular networks for pancreatic ductal adenocarcinoma progression. Cancer Cell Int. 2018;18:214.ArticleCASPubMedPubMed CentralGoogle Scholar
Miyake M, Morizawa Y, Hori S, Tatsumi Y, Onishi S, Owari T, Iida K, Onishi K, Gotoh D, Nakai Y, et al. Diagnostic and prognostic role of urinary collagens in primary human bladder cancer. Cancer Sci. 2017;108:2221–8.ArticleCASPubMedPubMed CentralGoogle Scholar
Nie XC, Wang JP, Zhu W, Xu XY, Xing YN, Yu M, Liu YP, Takano Y, Zheng HC. COL4A3 expression correlates with pathogenesis, pathologic behaviors, and prognosis of gastric carcinomas. Hum Pathol. 2013;44:77–86.ArticleCASPubMedGoogle Scholar
Souza P, Rizzardi F, Noleto G, Atanazio M, Bianchi O, Parra ER, Teodoro WR, Carrasco S, Velosa AP, Fernezlian S, et al. Refractory remodeling of the microenvironment by abnormal type V collagen, apoptosis, and immune response in non-small cell lung cancer. Hum Pathol. 2010;41:239–48.ArticleCASPubMedGoogle Scholar
Hou T, Tong C, Kazobinka G, Zhang W, Huang X, Huang Y, Zhang Y. Expression of COL6A1 predicts prognosis in cervical cancer patients. Am J Transl Res. 2016;8:2838–44.CASPubMedPubMed CentralGoogle Scholar
Zhang B, Zhang C, Yang X, Chen Y, Zhang H, Liu J, Wu Q. Cytoplasmic collagen XIalphaI as a prognostic biomarker in esophageal squamous cell carcinoma. Cancer Biol Ther. 2018;19:364–72.ArticleCASPubMedPubMed CentralGoogle Scholar
Moilanen JM, Kokkonen N, Loffek S, Vayrynen JP, Syvaniemi E, Hurskainen T, Makinen M, Klintrup K, Makela J, Sormunen R, et al. Collagen XVII expression correlates with the invasion and metastasis of colorectal cancer. Hum Pathol. 2015;46:434–42.ArticleCASPubMedGoogle Scholar
Galiger C, Loffek S, Stemmler MP, Kroeger JK, Mittapalli VR, Fauth L, Esser PR, Kern JS, Meiss F, Lassmann S, et al. Targeting of cell surface proteolysis of collagen XVII impedes squamous cell carcinoma progression. Mol Ther. 2018;26:17–30.ArticleCASPubMedGoogle Scholar
Liu CC, Lin SP, Hsu HS, Yang SH, Lin CH, Yang MH, Hung MC, Hung SC. Suspension survival mediated by PP2A-STAT3-Col XVII determines tumour initiation and metastasis in cancer stem cells. Nat Commun. 2016;7:11798.ArticleCASPubMedPubMed CentralGoogle Scholar
Spivey KA, Banyard J, Solis LM, Wistuba II, Barletta JA, Gandhi L, Feldman HA, Rodig SJ, Chirieac LR, Zetter BR. Collagen XXIII: a potential biomarker for the detection of primary and recurrent non-small cell lung cancer. Cancer Epidemiol Biomarkers Prev. 2010;19:1362–72.ArticleCASPubMedPubMed CentralGoogle Scholar
Wang Q, Yu J. MiR-129-5p suppresses gastric cancer cell invasion and proliferation by inhibiting COL1A1. Biochem Cell Biol. 2018;96:19–25.ArticleCASPubMedGoogle Scholar
Ivanovic RF, Viana NI, Morais DR, Silva IA, Leite KR, Pontes-Junior J, Inoue G, Nahas WC, Srougi M, Reis ST. miR-29b enhances prostate cancer cell invasion independently of MMP-2 expression. Cancer Cell Int. 2018;18:18.ArticlePubMedPubMed CentralCASGoogle Scholar
D’Aniello C, Cermola F, Palamidessi A, Wanderlingh LG, Gagliardi M, Migliaccio A, Varrone F, Casalino L, Matarazzo MR, De Cesare D, et al. collagen prolyl hydroxylation-dependent metabolic perturbation governs epigenetic remodeling and mesenchymal transition in pluripotent and cancer cells. Cancer Res. 2019;79:3235–50.ArticlePubMedGoogle Scholar
Atkinson A, Renziehausen A, Wang H, Lo Nigro C, Lattanzio L, Merlano M, Rao B, Weir L, Evans A, Matin R, et al. Collagen prolyl hydroxylases are bifunctional growth regulators in melanoma. J Invest Dermatol. 2019;139:1118–26.ArticleCASPubMedGoogle Scholar
Nagaraju GP, Park W, Wen J, Mahaseth H, Landry J, Farris AB, Willingham F, Sullivan PS, Proia DA, El-Hariry I, et al. Antiangiogenic effects of ganetespib in colorectal cancer mediated through inhibition of HIF-1alpha and STAT-3. Angiogenesis. 2013;16:903–17.ArticlePubMedGoogle Scholar
Dong H, Luo L, Zou M, Huang C, Wan X, Hu Y, Le Y, Zhao H, Li W, Zou F, Cai S. Blockade of extracellular heat shock protein 90alpha by 1G6-D7 attenuates pulmonary fibrosis through inhibiting ERK signaling. Am J Physiol Lung Cell Mol Physiol. 2017;313:L1006–15.ArticlePubMedCASGoogle Scholar
Bellaye PS, Burgy O, Causse S, Garrido C, Bonniaud P. Heat shock proteins in fibrosis and wound healing: good or evil? Pharmacol Ther. 2014;143:119–32.ArticleCASPubMedGoogle Scholar
Henke A, Franco OE, Stewart GD, Riddick AC, Katz E, Hayward SW, Thomson AA. reduced contractility and motility of prostatic cancer-associated fibroblasts after inhibition of heat shock protein 90. Cancers (Basel). 2016;8:E77.ArticleCASGoogle Scholar
Duarte BDP, Bonatto D. The heat shock protein 47 as a potential biomarker and a therapeutic agent in cancer research. J Cancer Res Clin Oncol. 2018;144:2319–28.ArticleCASPubMedGoogle Scholar
Zhu J, Xiong G, Fu H, Evers BM, Zhou BP, Xu R. Chaperone Hsp47 drives malignant growth and invasion by modulating an ECM gene network. Cancer Res. 2015;75:1580–91.ArticleCASPubMedPubMed CentralGoogle Scholar
Yao Q, Kou L, Tu Y, Zhu L. MMP-responsive ‘Smart’ Drug delivery and tumor targeting. Trends Pharmacol Sci. 2018;39:766–81.ArticleCASPubMedGoogle Scholar
Mohsen A, Collery P, Garnotel R, Brassart B, Etique N, Mohamed Sabry G, Elsherif Hassan R, Jeannesson P, Desmaele D, Morjani H. A new gallium complex inhibits tumor cell invasion and matrix metalloproteinase MMP-14 expression and activity. Metallomics. 2017;9:1176–84.ArticleCASPubMedGoogle Scholar
Trackman PC. Lysyl oxidase isoforms and potential therapeutic opportunities for fibrosis and cancer. Expert Opin Ther Targets. 2016;20:935–45.ArticleCASPubMedPubMed CentralGoogle Scholar
Wei MG, Sun W, He WM, Ni L, Yang YY. Ferulic acid attenuates TGF-beta1-induced renal cellular fibrosis in NRK-52E cells by inhibiting Smad/ILK/Snail pathway. Evid Based Complement Alternat Med. 2015;2015:619720.PubMedPubMed CentralGoogle Scholar
Cortes E, Lachowski D, Robinson B, Sarper M, Teppo JS, Thorpe SD, Lieberthal TJ, Iwamoto K, Lee DA, Okada-Hatakeyama M, et al. Tamoxifen mechanically reprograms the tumor microenvironment via HIF-1A and reduces cancer cell survival. EMBO Rep. 2019;20:e46557.PubMedGoogle Scholar
Szoka L, Karna E, Hlebowicz-Sarat K, Karaszewski J, Boryczka S, Palka JA. Acetylenic derivative of betulin induces apoptosis in endometrial adenocarcinoma cell line. Biomed Pharmacother. 2017;95:429–36.ArticleCASPubMedGoogle Scholar
Jiang H, Liu X, Knolhoff BL, Hegde S, Lee KB, Jiang H, Fields RC, Pachter JA, Lim KH, DeNardo DG. Development of resistance to FAK inhibition in pancreatic cancer is linked to stromal depletion. Gut. 2019. https://doi.org/10.1136/gutjnl-2018-317424.ArticlePubMedGoogle Scholar
Wang Z, Li J, Xiao W, Long J, Zhang H. The STAT3 inhibitor S3I-201 suppresses fibrogenesis and angiogenesis in liver fibrosis. Lab Invest. 2018;98:1600–13.ArticleCASPubMedGoogle Scholar
Zhang Q, Hou X, Evans BJ, VanBlaricom JL, Weroha SJ, Cliby WA: LY2157299 Monohydrate, a TGF-betaR1 Inhibitor, Suppresses Tumor Growth and Ascites Development in Ovarian Cancer. Cancers (Basel) 2018, 10.
Colak S, Ten Dijke P. Targeting TGF-beta signaling in cancer. Trends Cancer. 2017;3:56–71.ArticleCASPubMedGoogle Scholar
Averett C, Bhardwaj A, Arora S, Srivastava SK, Khan MA, Ahmad A, Singh S, Carter JE, Khushman M, Singh AP. Honokiol suppresses pancreatic tumor growth, metastasis and desmoplasia by interfering with tumor-stromal cross-talk. Carcinogenesis. 2016;37:1052–61.ArticleCASPubMedPubMed CentralGoogle Scholar
Wang T, Fu X, Jin T, Zhang L, Liu B, Wu Y, Xu F, Wang X, Ye K, Zhang W, Ye L. Aspirin targets P4HA2 through inhibiting NF-kappaB and LMCD1-AS1/let-7 g to inhibit tumour growth and collagen deposition in hepatocellular carcinoma. EBioMedicine. 2019;45:168–80.ArticlePubMedPubMed CentralGoogle Scholar
Khan S, Ebeling MC, Chauhan N, Thompson PA, Gara RK, Ganju A, Yallapu MM, Behrman SW, Zhao H, Zafar N, et al. Ormeloxifene suppresses desmoplasia and enhances sensitivity of gemcitabine in pancreatic cancer. Cancer Res. 2015;75:2292–304.ArticleCASPubMedPubMed CentralGoogle Scholar
Olsson PO, Gustafsson R, In’t Zandt R, Friman T, Maccarana M, Tykesson E, Oldberg A, Rubin K, Kalamajski S. The tyrosine kinase inhibitor imatinib augments extracellular fluid exchange and reduces average collagen fibril diameter in experimental carcinoma. Mol Cancer Ther. 2016;15:2455–64.ArticleCASPubMedGoogle Scholar
Grither WR, Longmore GD. Inhibition of tumor-microenvironment interaction and tumor invasion by small-molecule allosteric inhibitor of DDR2 extracellular domain. Proc Natl Acad Sci USA. 2018;115:E7786–94.ArticlePubMedCASPubMed CentralGoogle Scholar
Aguilera KY, Huang H, Du W, Hagopian MM, Wang Z, Hinz S, Hwang TH, Wang H, Fleming JB, Castrillon DH, et al. Inhibition of discoidin domain receptor 1 reduces collagen-mediated tumorigenicity in pancreatic ductal adenocarcinoma. Mol Cancer Ther. 2017;16:2473–85.ArticleCASPubMedPubMed CentralGoogle Scholar
Brandsma I, Fleuren EDG, Williamson CT, Lord CJ. Directing the use of DDR kinase inhibitors in cancer treatment. Expert Opin Investig Drugs. 2017;26:1341–55.ArticleCASPubMedPubMed CentralGoogle Scholar
Kobayashi M, Sawada K, Kimura T. Potential of integrin inhibitors for treating ovarian cancer: a literature review. Cancers (Basel). 2017;9:E83.ArticleCASGoogle Scholar
Compte M, Harwood SL, Munoz IG, Navarro R, Zonca M, Perez-Chacon G, Erce-Llamazares A, Merino N, Tapia-Galisteo A, Cuesta AM, et al. A tumor-targeted trimeric 4-1BB-agonistic antibody induces potent anti-tumor immunity without systemic toxicity. Nat Commun. 2018;9:4809.ArticlePubMedPubMed CentralCASGoogle Scholar
Chen X, Song E. Turning foes to friends: targeting cancer-associated fibroblasts. Nat Rev Drug Discov. 2019;18:99–115.ArticleCASPubMedGoogle Scholar
Alvarez R, Musteanu M, Garcia-Garcia E, Lopez-Casas PP, Megias D, Guerra C, Munoz M, Quijano Y, Cubillo A, Rodriguez-Pascual J, et al. Stromal disrupting effects of nab-paclitaxel in pancreatic cancer. Br J Cancer. 2013;109:926–33.ArticleCASPubMedPubMed CentralGoogle Scholar
Huang H, Brekken RA. The next wave of stroma-targeting therapy in pancreatic cancer. Cancer Res. 2019;79:328–30.ArticleCASPubMedGoogle Scholar
Sielaff CM, Mousa SA. Status and future directions in the management of pancreatic cancer: potential impact of nanotechnology. J Cancer Res Clin Oncol. 2018;144:1205–17.ArticleCASPubMedGoogle Scholar
Li Y, Hong J, Oh JE, Yoon AR, Yun CO. Potent antitumor effect of tumor microenvironment-targeted oncolytic adenovirus against desmoplastic pancreatic cancer. Int J Cancer. 2018;142:392–413.ArticleCASPubMedGoogle Scholar
Eriksson E, Milenova I, Wenthe J, Stahle M, Leja-Jarblad J, Ullenhag G, Dimberg A, Moreno R, Alemany R, Loskog A. Shaping the tumor stroma and sparking immune activation by CD40 and 4-1BB signaling induced by an armed oncolytic virus. Clin Cancer Res. 2017;23:5846–57.
Title: Bovine Collagen vs. Fish Collagen: Exploring the Key Differences and Making Informed Choices
Introduction:
Collagen is the most abundant protein in our bodies, providing structure to our skin, hair, nails, and connective tissues. As we age, our natural collagen production declines, leading many to turn to collagen supplements. Two common sources of collagen supplements are bovine (cow) collagen and fish collagen. In this blog post, we’ll delve into the differences between bovine and fish collagen, how to interpret product labels, recommended dosages, sources, efficacy, potential side effects, and toxicity concerns.
Understanding Bovine and Fish Collagen:
Bovine Collagen: Bovine collagen is derived from the skin, bones, and cartilage of cows. It is rich in Type I and Type III collagen, which are essential for skin, hair, and bone health. Bovine collagen is often used in powdered form or as an ingredient in various beauty and health products.
Fish Collagen: Fish collagen is sourced from the skin and scales of fish, such as cod or tilapia. It predominantly consists of Type I collagen, known for its benefits in improving skin elasticity and joint health. Fish collagen is popular among those with dietary restrictions or allergies to bovine sources.
How to Read the Label:
When shopping for collagen supplements, pay attention to the following details on the product label:
Source: Look for “bovine collagen” or “fish collagen” on the ingredient list to determine the source of collagen.
Type: Collagen supplements may specify the type of collagen present, such as Type I or Type III. This information can help you choose a product aligned with your specific health goals.
Dosage:
The recommended dosage for collagen supplements can vary depending on individual needs and the specific product. Generally, a daily dose of 2.5 to 10 grams is common. However, it’s essential to follow the manufacturer’s instructions and consult with a healthcare professional for personalized guidance.
Best Sources of Collagen:
Natural Sources: You can support your collagen production by consuming a balanced diet rich in foods like bone broth, chicken, fish, and lean meat. These foods contain nutrients that promote collagen synthesis, such as vitamin C, zinc, and amino acids.
Supplements: Collagen supplements can be a convenient way to boost your collagen intake, especially if your diet lacks collagen-rich foods. Choosing between bovine and fish collagen depends on your preferences, dietary restrictions, and specific health goals.
Efficacy and Amount:
Both bovine and fish collagen have been shown to promote skin elasticity, nail and hair growth, and joint health. The efficacy of each source depends on the type of collagen present and individual response. To maximize benefits, consistency in supplementation is key.
Potential Side Effects and Toxicity:
Collagen supplements are generally considered safe when taken as directed. However, some individuals may experience minor side effects such as digestive discomfort or allergic reactions. It’s essential to start with a low dose and gradually increase it if needed. If you have allergies or specific dietary restrictions, choose collagen supplements that align with your needs.
References:
Asserin J, Lati E, Shioya T, Prawitt J. The effect of oral collagen peptide supplementation on skin moisture and the dermal collagen network: evidence from an ex vivo model and randomized, placebo-controlled clinical trials. J Cosmet Dermatol. 2015;14(4):291-301.
Liu D, Nikoo M, Boran G, Zhou P. Regenstein. Collagen and gelatin. Annu Rev Food Sci Technol. 2015;6:527-557.
Schunck M, Zague V, Oesser S, Proksch E. Dietary supplementation with specific collagen peptides has a body mass index-dependent beneficial effect on cellulite morphology. J Med Food. 2015;18(12):1340-1348.
Conclusion:
Choosing between bovine and fish collagen ultimately comes down to your dietary preferences and specific health needs. Both sources offer valuable benefits, and collagen supplements can be a valuable addition to your wellness routine when used appropriately. By reading labels, understanding dosages, and considering individual factors, you can make informed decisions and support your collagen-related health goals effectively.